
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Дипломная работа: Эколого-физиологические особенности микроскопических грибов представителей рода Aspergillus, выделенных из разных местообитаний
Дипломная работа: Эколого-физиологические особенности микроскопических грибов представителей рода Aspergillus, выделенных из разных местообитаний
Эколого-физиологические особенности микроскопических грибов представителей рода Aspergillus, выделенных из разных местообитаний
Среди необходимых для жизни микроскопических грибов элементов углерод занимает совершенно особое положение. Около половины сухого веса гриба приходится на долю углерода. Этот элемент является постоянной составной частью протоплазмы, ферментов, клеточной оболочки и запасных питательных веществ. Поэтому соединения, содержащие углерод, играют важнейшую роль в питании грибов (Лилли, 1957).
Микроскопические грибы значительно различаются способностью усваивать разные соединения углерода и синтезировать из них составные части клетки. Некоторые виды удивительно всеядны и могут использовать для питания разнообразные соединения. С другой стороны, известно множество различных специализированных типов микромицетов, которые нуждаются в специфических соединениях. Нефть, газообразные углеводороды, парафины, даже резины, гудрон, капрон и многие другие синтетические материалы, а также пестициды после попадания в почву начинают разлагаться плесневыми грибами и бактериями. Практически не существует органических соединений, которые не усваивались бы микроорганизмами.
Ни один из видов грибов не способен усваивать все существующие в природе естественные органические соединения. Точно также и определенное органическое соединение не усваивается одинаково хорошо всеми видами грибов. Усваиваемость определяется составом, структурой и конфигурацией молекул органического соединения.
Любое органическое вещество вначале расщепляется до более простых соединений, а последние вовлекаются в тот или иной биосинтетический процесс. Возникающая при расщеплении энергия накапливается в АТФ или в других соединениях, имеющих макроэргические связи. Таким образом, органические субстраты обеспечивают как энергетическую, так и конструктивную стороны обмена плесневых грибов и бактерий. Следовательно, вопрос об углеродном питании и о влиянии источников углерода на развитие микроорганизмов является очень существенным при проведении любых микробиологических экспериментов (Лилли, 1957).
Целью данной работы является изучение эколого-физиологических особенностей микроскопических грибов представителей рода Aspergillus, выделенных из разных местообитаний.
В соответствии с поставленной целью задачами исследования являются:
1) определение способности штаммов использовать легкоразлагаемые источники углерода (сахара, многоатомные спирты, крахмал);
2) определение способности штаммов использовать трудноразлагаемые источники углерода (целлюлозу, нефть, гербициды и пестициды).
Практическая значимость данной работы заключается в том, что были впервые изучены пищевые предпочтения чистых культур микроскопических грибов, выделенных из различных местообитаний.
1. Особенности микроскопических грибов, выделенных из различных экониш
1.1 Экологические группы грибов
Гетеротрофный способ питания, характерный для грибов, обуславливает
то, что их развитие в большей степени зависит от субстрата, на котором они развиваются. В связи с высокими адаптивными возможностями грибы занимают самые разнообразные экологические ниши и играют значительную роль в биоценозах. При выделении экологических групп грибов учитывают как место их обитания, так и характер субстрата, который они используют для своей жизнедеятельности.
Почвенные грибы. В почве обитают представители практически всех отделов грибов. Одни из них находятся в ней постоянно – истинно почвенные грибы, другие попадают случайно с опадом листьев и растительными остатками, дождевыми и талыми водами, частицами пыли из воздуха и так далее. Случайно попавшие в почву грибы находятся в неактивном состоянии – в виде различного типа спороношений (конидий, телиоспор, склероциев) или в виде неактивного мицелия. Циклы развития этих грибов часто не связаны с почвой. Она служит только для их выживания и перезимовки (Гарибова, 2005).
В почве осуществляется основная функция грибов, как главных редуцентов в экосистемах суши. Представители почвенных сапротрофных микромицетов осуществляют важную экологическую функцию по разложению органических веществ, начиная от самых простых углеводов и кончая такими сложными биополимерами, как целлюлоза, хитин, лигнин и так далее. Сапротрофные микромицеты чрезвычайно разнообразны как в таксономическом отношении, так и по связи с разными субстратами и по приспособленности к факторам физической среды. Их образ жизни определяется характером обмена веществ, наличием разного рода приспособительных реакций, связанных с внешними условиями, устойчивостью к стрессовым воздействиям. Основной экологический фактор, определяющий разнообразие грибов, это тип субстрата как среда обитания.
В почвах одного типа, но под разными растительными ассоциациями, комплексы доминантных видов микромицетов могут быть резко различными. Например, в дерново-подзолистой почве под ельником доминирующими видами были Aspergillus pullulans, Chrysosporium sulfureum, Mortierella ramanniana и Verticillium terrestre, а в почве под березняком – A. pulviscula, C. herbarum, Penicillium daleae и Trichoderma koningii (Звягинцев, 2005).
Распределение грибов по профилю соответствует содержанию гумуса по горизонтам почвы. Для большинства почв характерно максимальное количество грибных зародышей в верхних гумусированных горизонтах и резкое снижение их числа и числа видов с глубиной. Большее число грибов в верхних горизонтах почв в первую очередь связано с большим содержанием органического вещества в этих горизонтах. С глубиной количество органического вещества убывает, одновременно снижается число грибных зародышей.
Установлено, что грибы могут развиваться в очень широких пределах значений pH, то есть как в сильнокислых, так и в щелочных почвах. Однако в связи с тем, что в кислых почвах грибы подвержены меньшей конкуренции со стороны бактерий, они развиваются там в большем числе, но большее разнообразие видов наблюдается в щелочных почвах. В видовом составе грибов кислых и щелочных почв существуют заметные различия, хотя ряд видов может быть общим. Только в кислых песчаных подзолах Англии выделены Penicillium janthinellum, P. frequentans, Mortierella ramanniana, только в щелочных карбонатных почвах выделены Fusarium sambucinum, P. luteum, Thielavia sp. (Мирчинк, 1988).
Из других факторов на грибное население почв значительно влияют температурные колебания в верхних слоях почвы. Большинство грибов – мезофилы, облигатные психрофилы описаны только среди дрожжевых грибов. Отношение к температуре определяет в значительной степени местообитание многих видов микромицетов или долю их популяций в сообществах в разные периоды года (Звягинцев, 2005).
К мезофилам относится гриб Penicillium frequenetes, обитающий в умеренной зоне, Cladosporium cladosporioides, обитающий как в умеренной зоне, так и далеко за ее пределами. Однако популяции этого вида, обитающие в разных зонах, имеют разные температурные границы роста и прорастания спор. Мезофилами также можно считать многие виды Penicillium, Fusarium, Mucor и многие другие.
К термофилам относятся многие организмы, выделяющиеся из саморазогревающихся торфов, такие, как Chaetomium thermophila, Sporotrichum thermophila, Talaromyces emersonii, Thermoascus aurantiacus, виды Humicola.
К термофилам и термотолерантным формам принадлежат многие виды Aspergillus. Так, популяция A. terreus, выделенная из пустынных почв Туркмении, имеет границы роста от 20 до 47 ○C при оптимуме 37 ○С (Мирчинк, 1988).
Водные грибы. Большая по объему группа грибов, связанная в своих местообитаниях с водой. Среди них виды – сапротрофы, живущие на растительных остатках, попавших в водную среду, паразиты водных животных, водорослей, высших водных растений. Причем водные грибы известны как в пресных, так и морских водоемах. К водным грибам относятся, например, сапролегниевые грибы, а также из многих таксономических групп.
Среди грибов, обитающих в водных экосистемах, выделяют две большие категории: постоянные обитатели водной среды и иммигранты, появляющиеся в водоемах периодически, с приуроченностью к определенным сезонам или без таковой.
Грибы первой группы (зооспоровые, водные гифомицеты) весь жизненный цикл проводят в воде и играют существенную роль в трофических сетях водоемов. Грибы второй группы – терригенные – неоднородны по своей приспособленности к существованию в водной среде, среди них есть неактивные формы, со временем исчезающие, и активные (Терехова, 2007).
В результате многолетних микологических исследований водных экосистем И.А. Дудка (1985) предложила разделять водные грибы на виды-резиденты, составляющие активное ядро гидромикобиоты, определяющее участие в функционировании гетеротрофного блока биогидроценоза, и транзитные виды.
На формирование состава микобиоты в водных экосистемах оказывают влияние степень трофности среды обитания, весь комплекс продукционно-биологических процессов, особенности гидродинамики и гидрохимии, наличие соответствующих организмов-хозяев для паразитирующих и субстратов для сапротрофных видов.
Наличие у многих микромицетов, постоянно выделяющихся из водоемов, способности образовывать широкий спектр внеклеточных ферментов, свидетельствует о возможности активно участвовать в деструкции органического материала в водной среде. Так, среди выделенных с погруженной в воду древесины и листьев грибов 18 видов были активными продуцентами амилазы, ксиланазы, целлюлазы, протеазы, пероксидазы и липазы; 5 видов разлагали хитин, 8 – выделяли тирозиназу и лакказу (Parton, 1988). Отмечают высокую целюлозолитическую активность у штаммов видов Trichoderma, Aspergillus niger и Penicillium miszynsky, выделенных из водных местообитаний (Hopper, 1995).
Микроскопические грибы могут прямо или косвенно влиять на активность и развитие других гидробионтов. Пути воздействия могут быть различными – от прямого паразитирования до опосредованного влияния через изменения среды обитания, путем выделения биологически активных метаболитов. Среди грибных экзометаболитов известно большое количество токсичных компонентов.
Г. Нейш и Г. Хьюз (1984) отмечали на рыбе Fusarium culmorum, Phoma herbarum. Л. Воронин (1987) сообщает о высокой частоте встречаемости на теле микромицетов из родов Cladosporium, Fusarium, Mortierella, Mucor, Penicillium, Phoma, Trichoderma.
Таким образом, многие виды терригенных микромицетов длительно и активно развиваются в воде, образуют спороношения, включаются в пищевые цепи водных экосистем, участвуя в деструкции органического материала в воде и донных отложениях.
При изучении процессов микробиологической деструкции веществ в водоемах наряду с бактериями должен учитываться вклад водных грибов: запасы грибной биомассы значимы, а высокая частота встречаемости, ферментативная и антибиотическая активность микроскопических грибов дают основания судить о грибах как важном элементе водных экосистем, способном изменять окружающую среду (Терехова, 2007).
Процесс формирования новых эколого-трофических групп грибов продолжается и в настоящее время. Возникающие в процессе деятельности человека новые материалы (стекло, пластмассы, бумага, различные, сложные по составу промышленные материалы, нефтепродукты и так далее), приводят к тому, что грибы адаптируются к этим специфическим субстратам, осваивают их и, таким образом, формируют новые эколого–трофические группы (Гарибова, 2005).
1.2 Влияние экологических условий на рост микромицетов
1.2.1 Особенности роста микроскопических грибов в стандартных условиях культивирования
А.Е. Ивановой (1999) было исследовано формирование микроколоний из двух типов колониеобразующих единиц (при размножении спорами и фрагментами мицелия) в стандартных для почвенно–микробиологических анализов условиях – на среде Чапека при 25 ○С после взбалтывания на качалке.
Было установлено, что в зависимости от вида гриба и типа КОЕ наличие и величина лаг-фазы могут различаться. Так, у гриба Mucor hiemalis длительность лаг-фазы при росте мицелия из фрагментов гиф разной длины была меньше, чем при росте из спор. А при росте мицелия из фрагментов гиф Alternaria alternatа и Penicillium spinulosum лаг-фаза в данных условиях вовсе не наблюдалось. В то же время при прорастании спор A. alternatа лаг-фаза тоже не отмечена, а прорастание спор P. spinulosum наступало лишь через определенный интервал времени.
Длительность экспоненциальной фазы была короче при развитии грибных микроколоний из фрагментов мицелия, чем из спор. Чем больше была начальная величина растущих фрагментов, тем короче были сроки прохождения экспоненциональной фазы роста. Удельная скорость роста в экспоненциональной фазе при развитии мицелия из фрагментов гиф M. hiemalis и P. spinulosum была меньше, чем при развитии из спор. Для этих видов характерны мелкие споры, а фрагменты имеют в несколько раз больший первоначальный объем гиф. При формировании микроколоний A. alternatа скорости экспоненциального роста мицелия из разных КОЕ существенно не отличались, возможно, благодаря малой разнице в размерах конидий A. alternatа (d=10–30 мкм) и жизнеспособных мелких фрагментов.
Ветвление мицелия, растущего из разных КОЕ, на первых этапах формирования грибных микроколоний также может отличаться. В экспоненциальной фазе роста в микроколониях, растущих из спор, единица гифального роста (ЕГР) часто была выше (то есть ветвление реже), чем в микроколониях, растущих из фрагментов гиф.
Линейный рост при развитии колоний из фрагментов гиф наступал раньше. Далее в этой фазе скорости мицелия из фрагментов гиф и спор грибов не различались; величина ЕГР стабилизировалась и не зависела от типа исходных КОЕ. То есть, в линейной фазе роста все различия между микроколониями, сформированными из разных КОЕ, нивелировались (Иванова, 1999).
Особый интерес заслуживает характер ритмических колебаний роста исследуемых культур.
Биологические ритмы присущи всем уровням живой материи – от молекулярных и субклеточных структур до биосферы, что обеспечивает единство живой и неживой природы. Все они отражают процессы регулирования функций организма (Романов, 1980).
Биохимический механизм циркадных ритмов ввязывают с обменом нуклеиновых кислот (Шаркова, 1971). Имеющиеся данные указывают на то, что с усилением ритма инициируется обмен РНК. Биоритмы митоспоровых грибов отражают неограниченно восстанавливаемый адаптивный онтогенез, формирующийся на основе сигналов окружающей среды, генетический механизм которого базируется на высокой способности грибов к формированию многоядерности и ядерного дуализма (Беккер, 1983).
Характер и продолжительность ростовых ритмов, так же как и радиальная скорость роста, зависят от систематического положения гриба, его возраста (возраста культуры), концентрации в среде основных источников питания и энергии, а также других экологических факторов (Романов, 1980; Бухало, 1988).
1.2.2 Влияние экологических условий на жизнеспособность мицелия микроскопических грибов
Было проведено исследование А.Е. Ивановой (1999) с целью определения жизнеспособности грибного мицелия разной длины в различных экологических условиях среды. Так, увеличение концентрации органического вещества (сахарозы) в интервале 0–20 г./л было благоприятно для фрагментов Mucor hiemalis, способность к росту у которых возрастала. Высокая концентрация сахарозы (100 г./л), наоборот, подавляла рост крупных фрагментов M. hiemalis. Число жизнеспособных фрагментов мицелия разной длины Penicillium spinulosum больше при низких концентрациях сахарозы, а ее высокое содержание подавляет рост мелких (30–60 мкм) фрагментов. При высоком содержании сахарозы увеличивается доля растущих после взбалтывания на качалке коротких фрагментов (20–100 мкм, или 3–6 клеток) Alternaria alternatа, а жизнеспособность крупных фрагментов гиф (> 130 мкм, или > 9 клеток) не изменялась. Однако, после обработки ультразвуком высокий уровень сахарозы (100 г./л) подавлял рост фрагментов A. alternatа любой длины.
При низких температурах у всех исследованных видов грибов (Alternaria alternata, Mucor hiemalis, Penicillium spinulosum) было отмечено значительное снижение способности к росту. Наиболее чувствительным оказался вид P. spinulosum, для которого 4 ○С это нижний температурный предел роста: при 4 ○С даже крупные фрагменты практически не растут. Мелкие фрагменты M. hiemalis тоже теряли способность к росту при 4 ○С. В несколько (4–6) раз уменьшалась жизнеспособность фрагментов A. alternatа.
Максимальная способность к росту мелких фрагментов M. hiemalis отмечалась при 25, 30 ○С. Жизнеспособность фрагментов A. alternatа практически не изменялась при 20, 25, 30 ○С. Мелкие и крупные фрагменты P. spinulosum наибольшую способность к росту проявляли при 20 ○С, при увеличении температуры до 30 ○С их жизнеспособность снижалась.
Изменение кислотности среды не оказывало существенного влияния на жизнеспособность фрагментов M. hiemalis и A. alternatа. Число способных к росту фрагментов P. spinulosum было наибольшим в нейтральных условиях среды, уменьшение pH от 7,0 до 3,5 приводило к снижению жизнеспособности всех фрагментов P. spinulosum, а мелкие фрагменты в кислых условиях не росли вовсе. С увеличением кислотности наблюдается подавление роста фрагментов мицелия. В варианте с максимальной кислотностью среды (pH 3,0) рост коротких (20–50 мкм) и средней длины (51–100 мкм) фрагментов отсутствовал практически полностью. Наиболее стабильный и сбалансированный рост фрагментов различной длины отмечается в вариантах с нейтральной и слабощелочной реакцией среды – 7,0 и 8,0 pH. При этом активный рост наблюдается и у коротких, и у длинных фрагментов (Григорьев, 2004).
Загрязнение возрастающими дозами тяжелого металла – кадмия оказывало негативное влияние на способность к росту всех фрагментов P. spinulosum. Напротив, присутствие кадмия вызывало повышение жизнеспособности фрагментов разной длины мицелия A. alternatа и малых (85 – 140 мкм) фрагментов M. hiemalis (Иванова, 1999).
1.3 Особенности использования микроскопическими грибами источников углерода
1.3.1 Разложение легкоусвояемых органических веществ
Важнейшая, быстрее всего усвояемая пища плесневых грибов состоит из моносахаридов и других низкомолекулярных водорастворимых соединений углерода, которые могут непосредственно поглощаться протопластом. Почти все организмы ассимилируют простые сахара и аналогичные им молекулы одинаково, однако грибы, конкурируя за эти питательные вещества, обладают некоторыми существенными преимуществами. Зачатки «сахарных грибов» присутствуют практически повсюду. Как только какой–либо живой или мертвый органический субстрат основательно увлажняется, возникает водный раствор, содержащий по крайней мере следы питательных веществ. Тотчас же там развиваются талломы «подходящих» грибов, быстро образуются их новые вегетативные единицы, и стремительно размножающаяся популяция полностью берет на себя использование данного источника пищи. За подобные субстраты с грибами конкурируют бактерии; получит ли преимущество кто-то из них или они будут сосуществовать относительно «равноправно», зависит от обстоятельств, и общего правила здесь вывести невозможно.
В отсутствие источников азота некоторые грибы окисляют глюкозу до глюконовой кислоты. При этом pH падает ниже 2,0, и обычные бактерии уже не могут размножаться, однако сами грибы при последующем поступлении азота способны утилизовать глюконовую кислоту. Примеров, объясняющих превосходство грибов над бактериями особенностями первичного обмена веществ, немного. Что касается превращений низкомолекулярных органических соединений, для грибов специфичны определенные пути разложения сахаров.
Разложение сахаров. В клетку часто проникают моносахариды – продукты внеклеточного разложения полисахаридов; ди- и олигосахариды также поглощаются из окружающей среды и включаются в метаболизм. Необходимые для этого ферменты либо широко распространены (мальтоза, сахароза и так далее), либо обнаружены у более или менее многих представителей грибов (Lodder, 1970; Barnett, 1979).
Наиболее обычный источник углерода – глюкоза. Полное разложение одного ее моля дает 675 ккал энергии. Другие гексозы (глюкоза – не обязательно) включаются в универсальный процесс разложения только после фосфорилирования; этим же путем идут продукты расщепления внеклеточных полисахаридов, запасных и входящих в состав клеточной стенки макромолекул.
Первая реакция разложения гексоз протекает с использованием энергии по общей формуле:
Гексоза + АТФ → Гексозофосфат +АДФ.
При этом с участием фермента гексокиназы из глюкозы, фруктозы и маннита возникают соответствующие гексозо-6-фосфаты, а из галактозы под действием галактокиназы галактозо-1-фосфат, который затем изомеризуется. Разложение сахаров протекает в грибной клетке следующими основными путями.
Фруктозодифосфатный путь (ФДФ, гликолиз, путь Эмбдена-Мейргофа-Парнаса) может вести к полному окислению, неполностью окисленным конечным продуктам или ответвляться в сторону образования сырья для биосинтеза. Гликолитические реакции в клетке начинаются с фосфорилирования глюкозы (в форме фосфатов сахара более рекционноспособны). При трансформации глюкозы в пировиноградную кислоту по пути Эмбдена-Мейргофа-Парнаса выделяется свободная энергия, достаточная для образования четырех молекул АТФ. Однако две из них требуются для превращения глюкозы в фруктозо – 1,6-дифосфат, и только две молекулы АТФ остаются для процессов синтеза.
Пентозофосфатный путь (ПФ) либо поставляет промежуточные продукты для последующего биосинтеза, в том числе нуклеотидов, либо продолжается по типу ФДФ.
При разложении через 2-кето-3-дезокси-6-фосфоглюконовую кислоту (КДФГ, путь Энтнера-Дудорова =ЭД) продукты расщепления КДФГ, образующейся путем дегидратации 6-фосфоглюконовой кислоты (глицеральдегидфосфат, пировиноградная кислота; быстро и непосредственно попадают в систему гликолиза).
С помощью глюкооксидазы (ГО) некоторые виды Aspergillus и Penicillium окисляют непосредственно глюкозу до глюконовой кислоты, которая выделяется в среду или включается в ПФ; возникающая при этом ядовитая для всех организмов перекись водорода ферментативно разрушается.
В глиоксилатном цикле (ГЦ) – побочном пути цикла лимонной кислоты (ЦЛК) изолимонная кислота, возникающая из ацетил – кофермента А и щавелевоуксусной кислоты, превращается в янтарную и глиоксиловую кислоты; последняя, реагируя с ацетил – коферментом А, дает яблочную кислоту, позволяющую продолжаться ЦЛК. Недостаток субстрата для ЦЛК может возникать, например, из–за расходования α – кетоглутаровой кислоты для синтеза аминокислот, тогда глиоксилатный путь заменяет отсутствующие промежуточные звенья (реакции пополнения, анаплеротические последовательности). Глюкоза и продукты ее разложения стимулируют нормальное протекание ЦЛК и подавляют ГЦ, который может активироваться присутствием в среде ацетата или глицина.
Полное окисление. С помощью дыхательных ферментов процесс соединения водорода с кислородом, дающий энергию почти всем организмам, подразделяется на мелкие этапы с незначительными различиями в энергосодержании исходных веществ и продуктов («биологический взрыв гремучего газа»). В ходе этих отдельных реакций, в частности, регенерируется АТФ. Ферментные системы различных организмов, несмотря на существенные общие черты неодинаковы. Так, у оомицетов отсутствует цитохром С1, свойственный грибам и растениям, а у одного из представителей рода Aspergillus отмечен цитохром, не отравляемый цианидом – (аналогичный В-цитохрому растений).
Доступность и использование различных путей разложения углеводов. То, какой путь задействован, зависит от организма, среды и состояния клетки, например от активности ее ферментов. Для определения этого количественно оценивают превращения субстрата соответствующими ключевыми ферментами.
Полиолы. Многоатомные спирты (полиолы), например маннит, рибит, глицерин, – результат окисления глюкозо-фосфата или соответствующих предшественников, конкурентного глюконеогенезу, спиртовому брожению или полному окислению в ЦЛК, а также синтезу макромолекул или другим реакциям с использованием АТФ и восстановлением НАДФ. Возможно, полиолы вместе с трегалозой служат у грибов формой транспорта углерода в гифах; они способны регулировать восстановительную силу, энергоснабжение, осмотические условия, содержание запасных веществ и рост. Некоторые авторы считают состав полиолов у грибов важным таксономическим признаком: у хитридиомицетов, аскомицетов, базидиомицетов и дейтеромицетов преобладает маннит, у зигомицетов его нет или же он не относится к главным компонентам; в целом у грибоподобных протистов полиолов меньше, чем у настоящих грибов. Наряду с такими наиболее частыми многоатомными спиртами, как глицерин (почти у всех) и манит (у всех грибов, кроме ряда зигомицетов), обнаружены также эритрит, рибит и арабит, известные и у водорослей. Концентрация рибита у Mucorales (Zygomycetes) зависит от питания (рост при потреблении рибозы). Арабит в крови человека указывает на грибную инфекцию (например, поражение Candida albicans); у здоровых людей он отсутствует. Здесь также выделяются оомицеты, у которых не обнаружено никаких полиолов; некоторые другие группы низших грибов (грибоподобных протистов) до сих пор изучены в этом плане недостаточно (Мюллер, 1995).
1.3.2 Разложение трудноразлагаемых веществ
Плесневые грибы в качестве источника углерода могут использовать такие трудноразлагаемые вещества, как целлюлоза, крахмал, лигнин, пектиновые вещества, нефть, пестициды.
Разложение целлюлозы. Основными источниками целлюлозы для грибов в природных условиях служат древесина и различные растительные остатки. Среди микроскопических грибов, разлагающих целлюлозу известны представители следующих родов: Aspergillus, Coriolus, Eupenicillium, Fusarium, Penicillium, Sporotrichum, Trichoderma, Verticillium. Но только некоторые из них продуцируют полные внеклеточные целлюлолитические системы (эндо- и экзоглюканазы β-глюкозидазу). Среди них Trichoderma viride, T. reesei, T. koningii, Penicillium funiculosum, Fusarium solani. Для культуральной жидкости большинства других грибов этой группы характерно отсутствие экзоглюканазы, то есть эти грибы могут деградировать более аморфные формы целлюлозы.
Деградация высокоупорядоченной формы целлюлозы осуществляется благодаря синергическому действию комплекса целлюлолитических ферментов. При любой комбинации экзо- и эндоглюканаз Trichoderma koningii, Fusarium solani, Penicillium и Funiculosum отмечается выраженный синергизм. Однако синергизм между экзоглюканазами этих грибов и эндоглюканазами грибов, не продуцирующих экзоглюканазу (Myrothecium verrucaria), не выявлен. Нет также синергизма между экзоглюканазами грибов и эндоглюканазами рубцовых бактерий. Последнее указывает на существенные различия целлюлолитических систем грибов и бактерий (Марьиновская, 2006).
Целлюлоза является линейным полимером d-глюкозы. Остатки глюкозы в молекуле клетчатки, как и в молекуле целлобиозы связаны β-гликозидной связью. Поэтому клетчатку можно рассматривать как полимер целлобиозы. Нормэн и Фуллер (1942) считают, что большинство грибов способно усваивать клетчатку. Несмотря на то, что использование клетчатки грибами имеет большое значение в круговороте веществ в природе, процесс этот изучен далеко не полно.
Обычно считают, что первым этапом использования целлюлозы грибами является ее гидролиз. Гидролиз целлюлозы можно схематически представить следующим образом: целлюлоза → целлодекстрины → целлотетроза → целлобиоза → d-глюкоза. Известен штамм Aspergillus oryzae, выделяющий целлюлазу и целлобиозу (Лилли, 1957).
Разложение крахмала. Как и целлюлоза, крахмал является полимером d-глюкозы. Остатки глюкозы в его молекуле соединены между собой α-гликозидной связью, поэтому основной структурной единицей молекулы крахмала, как и молекулы гликогена, следует считать мальтозу. Зеленые растения синтезируют крахмал, животные и грибы образуют гликоген. Ферментативный гидролиз крахмала может быть схематически представлен следующим образом: крахмал→декстрины→мальтоза→d-глюкоза. Декстрины, имеющие разветвленную углеродную цепочку, лишь частично гидролизуются амилазой. Декстрины с неразветвленной углеродной цепочкой полностью превращаются в мальтозу (Мирчинк, 1988).
Крахмал нерастворим в воде. Лишь грибы, образующие амилазу, обладают способностью усваивать крахмал. Существует немало грибов, неспособных развиваться на средах с крахмалом, однако большинство из них может усваивать этот полисахарид. Было установлено, что 26 различных изученных видов и штаммов оомицетов из числа сапролегниевых усваивали как крахмал, так и продукты его гидролиза (декстрины, мальтозу и глюкозу), но не были способны ассимилировать 13 других источников углерода, включая сюда и фруктозу. Позднее А.С. Марголин (1942) показал, что 19 из 21 вида грибов, усваивающих мальтозу, обладали также способностью использовать и декстрин (Лилли, 1957).
Разложение пектиновых веществ. Среди грибов имеются активные разлагатели пектина, который является существенным компонентом растительного опада. Пектин образует в растениях межклеточное вещество, из которого состоят так называемые срединные пластинки, соединяющие между собой отдельные клетки растения. Они придают тканям прочность. Пектин представляет собой высокомолекулярное соединение углеводной природы – полисахарид, в котором метоксилированные остатки галактуроновой кислоты связаны между собой β-1,4-глюкозидными связями.
Многие грибы образуют пектинолитические ферменты. Высокая пектинолитическая активность обнаружена у некоторых эпифитных грибов, главным образом Aureobasidium pullulans и видов Cladosporium. Пектинолитические грибы занимают значительное место среди типичных представителей лесной подстилки – это виды родов Cladosporium, Alternaria, Aposphaeria, Penicillium, фитопатогенные грибы родов Fusarium, Verticillium, Botrytis cinerea, Sclerotinia sclerotiorum.
Ферментативное разрушение пектиновых веществ в растениях имеет значение в патогенезе некоторых заболеваний. Фитопатогенные грибы разрушают пектин срединной пластинкой и пектаты в первичных клеточных оболочках, что приводит к изменению их физико-химических свойств и создает условия для внедрения паразита, а также в результате действия пектинэстеразы образуются вещества – полигалактурониды, способные закупоривать сосуды, что в конечном итоге приводит к увяданию растений.
Существенное значение разрушения пектиновых веществ грибами имеет при разложении растительного опада. Практическое использование пектиназ грибов – применение в пищевой промышленности при приготовлении фруктовых соков для их осветления, а также при мочке льна (Мирчинк, 1988).
Участвуя в разложении многих углеродсодержащих веществ растительного опада и древесины в первую очередь трудноразлагаемых полимерных соединений, где грибам принадлежит ведущая роль, они занимают значительное место в круговороте углерода, являясь поставщиками СО2 в атмосферу.
Среди грибов есть организмы, разлагающие жиры и воска, входящие в состав растительных и животных тканей. Это определяется наличием у них ферментов липаз. Наибольшей активностью липолитических ферментов обладают виды Mucor lipolyticus, Rhizopus nigricans, Aspergillus niger, Penicillium verrusum, Penicillium roquefortii. Многие выделены с поверхности растений, являясь эпифитами и способны разлагать также восковые налеты на поверхности растений.
Известна также способность грибов разлагать как алифатические, так и ароматические углеводороды. В этом отношении наибольшей активностью характеризуются грибы рода Aspergillus (Мирчинк, 1988).
Разрушение грибами нефтепродуктов. В последние десятилетия в связи с возродившимся интересом к процессам микробного превращения углеводородов были обнаружены мицелиальные грибы, деятельность которых приводит к деструкции нефти и ее производных. В настоящее время доказано, что утилизировать нефтепродукты, в том числе различные топлива, во время хранения и транспортировки способны многие виды грибов и бактерий (Андреюк, 1980).
Нефтепродукты как среда обитания грибов характеризуются рядом особенностей: 1) содержат большое количество сравнительно доступного углерода и минимальное – азота при почти недоступном пространственном расположении его в молекуле; 2) в них почти отсутствует доступная активная вода. Это оказывает существенное влияние на синтез de novo грибной клетки.
Вопросы необходимого соотношения C:N у грибов при росте на нефтепродуктах в биохимическом аспекте исследованы еще мало и уровень этих данных уже не отвечает современным представлениям о возможностях грибной клетки. Очевидно, здесь имеет место не только типичный гетеротрофный процесс, но также определенное подобие хемотрофии и автотрофии, причем стадии роста отличаются и специфичны по способности к разным типам трофики. Особенно это проявляется в период формирования репродуктивных структур (Ниязова, 1982; Бабьева, 1983). Спецификой роста грибов на нефтепродуктах является их способность распространяться на поверхности, то есть возможность использовать при этом активную воду из воздуха, а также расти в толще нефтепродуктов, то есть ограничивать свои потребности в воде за счет активной воды самих нефтепродуктов (Евдокимова 1982).
Рост грибов (кладоспориев, пенициллиев, аспергиллов и некоторых других видов и штаммов) в разных нефтепродуктах характеризуется различным типом размещения мицелиальной пленки. Наиболее типичный – на разделе фаз, однако чаще всего наблюдается еще и глубинный рост, при котором развивается не только в толще жидкости – до 20 см. Причем интересно, что рост этих штаммов при определенном соотношении нефтепродуктов и воды мало зависит от высоты слоев смеси, а также воздуха в надсубстратном пространстве. Это свидетельствует о большой возможности мицелиальных грибов выдерживать жесткие условия и приспосабливаться к потреблению необходимых для метаболизма веществ не совсем обычными биохимическими и физиологическими путями.
В настоящее время установлено, что способность окислять углеводороды нефти не является специфической чертой отдельных видов грибов. Это не редкая их особенность, а одна из физиологических функций. Однако, несмотря на большое сходство химических и физических свойств фракций нефтепродуктов, у большинства видов грибов четко проявляется избирательное отношение к их утилизации (Бабьева, 1983).
Разрушение полимерных материалов. Синтез полимеров и создание на их основе материалов, обладающих повышенной стойкостью к факторам окружающей среды и воздействию различных организмов, привел к обострению экологической обстановки из-за накопления больших объемов отходов, содержащих эти соединения в разных отраслях промышленности. В последние десятилетия во многих странах уделяется большое внимание созданию полимерных материалов и их модификаций, утилизация которых возможна под воздействием микробиоты. В качестве добавок к пластификаторам исследователи используют природные компоненты такие, как крахмал, производные целлюлозы, протеин, хитозан и так далее. На основе этих композитных полимеров ряд фирм выпускает пластики для производства изделий разового пользования, упаковки пищевых продуктов, плоских пленок и так далее, которые обладают способностью к биодеградации при компостировании и так далее (Власова, 2001; Фомин, 2001). Состав микроорганизмов, контаминирующих техногенные материалы и способных вызывать их биодеградацию, очень разнообразен как в таксономическом отношении, так и по их физиолого-биохимической активности. Среди них ведущее место занимают представители дейтеромицетов, способные развиваться на обширном сортименте материалов, содержащих соединения как природного происхождения, так и искусственного синтеза (Биоповреждения, 1987; Коваль, 1989).
Было проведено исследование О.В. Сычуговой с соавторами (2003) с целью изучения возможности роста и развития видов микромицетов на композиции пленочного сополимера этилена и винилацетата с термопластичным крахмалом.
В процессе данного исследования оценка возможности потребления разных форм крахмала тест культурами показала, что они способны утилизировать данный источник углеродного питания. Однако динамика роста видов на разных средах при одинаковых условиях инкубации, при одной и той же навеске крахмала не одинакова, что особенно четко проявляется на 4–10-е сутки. Выявляется и некоторая разница в темпе роста видов грибов на нативном и растворимом крахмале разного происхождения, а также на средах Чапека и Гетченсона, взятых в качестве контроля (Сычугова, 2003).
Изменения морфологических признаков и образования новых структур у тест – культур на модифицированных средах при замене сахарозы на крахмал и росте на полимере не отмечаются, и они сопоставимы с параметрами, приведенными в определителях (Пидопличко, 1953; Ellis, 1971; Watanabe, 2000). Отличия выявлены у них только в темпах формирования морфологических структур. Хотя в литературе и приведены данные о влиянии субстрата на появление новых морфологических структур у грибов (Богомолова, 2001), однако, вероятнее всего, онтогенез и темпы развития определяются геномом вида, реализация программы которого зависит от влияния различных факторов (Шевцова, 1987; Долгова, 1997).
На поверхности пленки, содержащей термопластичный крахмал фиксируются сформированные пучки конидиеносцев Aspergillus niger, Paecilomyces variotii, Penicillium funiculosum, Chaetomium globosum, Trichoderma viride. Другие виды из взятого набора тест-культур не растут на данном субстрате или формируют слабое спороношение и в более поздние сроки.
Рост мицелия и формирование спороношения на композиции пленочного сополимера дает основание предполагать участие некоторых видов грибов в биодеструкции полимера из сополимеров этилена и винилацетата (СЭВА) с добавлением термопластичного крахмала (ТПК). Это дает возможность при последующих пересевах на смесях СЭВА и ТПК отобрать наиболее активные виды и их штаммы для разработки биотехнологии по его утилизации (Сычугова, 2003).
Таким образом, микроскопические грибы могут использовать в качестве источников углерода разнообразные органические вещества, тем самым являясь важными деструкторами различных природных материалов: целлюлозы, крахмала, лигнина, гемицеллюлозы, жиров, углеводородов, а также синтетических материалов, таких как пластики, пленки, упаковки пищевых продуктов и так далее.
2. Объекты и методы исследований
2.1 Объекты исследований
В качестве объектов исследования были выбраны различные виды микроскопических грибов рода Aspergillus: A. niger, A. ustus, A. terreus, A. flavus, A. fumigatus. Культуры были взяты из коллекции культур микроорганизмов кафедры «Прикладная биология и микробиология» АГТУ. Данные виды микромицетов были выделены из основных типов почв Астраханской области.
В качестве источника углерода были выбраны из легкоусвояемых – сахара: сахароза, арабиноза, ксилоза, галактоза, лактоза и мальтоза; многоатомные спирты: глицерин, сорбит, маннит; а также из полисахаридов – крахмал. Все эти источники были взяты из коллекции химических реактивов кафедры. Из трудноразлагаемых источников углерода были использованы: нефть, целлюлоза, гербицид и пестицид. Целлюлозу добавляли в среду в виде мелко нарезанной фильтровальной бумаги. Для определения способности микроскопических грибов рода Aspergillus использовать в качестве единственного источника углерода гербициды в данной работе использовали гербицид «Пиримифос-метил» или О, О-диметил-О – (2-диэтиламино-6-метилпиримидил-4) тиофосфат – это жидкость соломенного цвета, практически не растворим в воде, хорошо растворим во многих органических растворителях, неустойчив в кислой и щелочной средах, является эффективным средством для борьбы со многими насекомыми и клещами. Для определения способности микромицетов рода Aspergillus использовать пестициды в качестве источника углерода применяли дуст. Сахара, крахмал и многоатомные спирты добавлялись в среды в количестве 30 г./л, нефть 30 мл/л, целлюлоза – 10 г./л, гербицид – 0,25 мл/л и пестицид – 0,001 г./л.
2.2 Принципы составления питательных сред для грибов. Основные принципы композиции сред
При составлении питательных сред для грибов обычно пользуются результатами предварительных исследований по выяснению значения для роста и развития изучаемого объекта концентрации отдельных компонентов (источников углерода, азота, зольных веществ и витаминов).
Основными правилами, которых придерживаются при составлении сред, способствующих росту грибов, являются следующие:
1) целесообразно применять отдельные источники углерода и азота;
2) концентрация вещества, служащего источником азотного питания, должна сильно уступать концентрации вещества – источника углерода (примерно в 10 – 15 раз).
Хотя среды натурального происхождения более благоприятны для роста большинства грибов, однако при физиологических экспериментах по изучению развития и обмена у грибов использовать их нежелательно, так как их состав непостоянен. Для этих экспериментов обычно используются синтетические среды. Одной из первых синтетических сред для грибов (культур видов аспергиллов и пенициллов) была среда Роллена следующего состава (г/л):
сахароза 72 (около 5%);
винная кислота – 4 (для подкисления среды);
фосфорнокислый аммоний – 4;
углекислый калий – 0,6;
сернокислый аммоний – 0,25;
сернокислый цинк – 0,07;
сернокислое железо – 0,07;
кремнекислый калий – 0,07;
дистиллированная вода – 1500 мл.
Более простой состав имеет среда Чапека (г/л):
сахароза – 30;
NaNO3 – 2;
MgSO4 – 0,5;
FeSO4 – 0,01;
KH2PO4 – 1;
KCl – 0,5;
дистиллированная вода – 1000 мл.
Для многих грибов среды являются неполноценными. Некоторые грибы (особенно витаминозависимые) растут на них плохо или совсем не растут. В таких случаях, если потребность данного организма в витаминах не изучена, необходимо добавлять в среду экстракты растительных или животных тканей, выбирая их исходя из экологии данного гриба. Например, в среды для выращивания грибов – дереворазрушителей добавляют опилки из древесины той породы дерева, которую они поражают в природе, для выращивания грибов – паразитов растений – ткани растения – хозяина; выращивание грибов – копрофилов производится на средах с навозным экстрактом (Беккер, 1983).
2.3 Приготовление питательных сред
Посуда для приготовления сред не должна содержать посторонних веществ, например щелочей, выделяемых некоторыми сортами стекла. Перед употреблением посуду тщательно мыли, полоскали и высушивали. Среды варили в стеклянных колбах объемом 250 мл. Каждой среды готовили объемом по 100 мл, рассчитанной на 5 культур исследуемых штаммов. После варки среды стерилизовали в автоклаве при 0,5 атм. и 120 ○С в течение 20 минут. После стерилизации и добавления соответствующих источников углеродного питания среды разливали в стерильные чашки Петри по 20 мл. Предварительно в чашки вносят по 1 капле молочной кислоты для подкисления среды, которое необходимо, чтобы убить бактерии.
2.4 Определение способности плесневых грибов использовать соединения углерода
Микромицеты характеризуются неодинаковой способностью использовать различные соединения углерода для конструктивного и энергетического метаболизма. Чтобы выяснить возможность роста гриба за счет тех или иных углеродсодержащих веществ, их высевают на синтетические среды, содержащие в качестве единственного источника углерода различные моно-, ди- и полисахариды, многоатомные спирты, органические кислоты, углеводороды.
В данной работе определение особенностей роста грибов на различных источниках углеродного питания проводилось путем поверхностного посева исследуемых штаммов на среду Чапека с различными источниками углерода и среду Частухина (для целлюлозоразрушающих грибов). Посев культур осуществляли уколом в центр чашки Петри. Время культивирования составляло 14 суток, температура культивирования 25±2 ○С. Значение применяемых питательных сред для процессов роста грибов оценивалось методом измерения радиальной скорости роста путем периодического замера диаметра колоний грибов (через каждые 48 часов), растущих на чашках Петри.
Для определения способности микромицетов использовать различные источники углерода применяют и другие методики.
Например, многие микромицеты могут использовать в качестве единственного источника углерода органические кислоты. Для определения способности расти на средах с органическими кислотами рекомендуется плотная среда состава (г/л): (NH4)2HPO4 – 0,5; MgSO4*7H2O – 0,2; NaCl – 0,1; агар – 15,0; органическая кислота в виде соли Na или К – 2,0; pH 6,8. До стерилизации к среде добавляют 20 мл 0,04% водного раствора индикатора метилрот, который в интервале pH 6,8 – 8,4 изменяет окраску от желтой к красной. Среду разливают в пробирки и стерилизуют при 1 атм. Посев проводят уколом. Продолжительность культивирования от 2 до 14 суток в зависимости от скорости роста микроорганизмов. О потреблении органических кислот свидетельствует рост по уколу и изменение кислотности среды в щелочную сторону, что отчетливо заметно по цвету индикатора.
Некоторые грибы способны использовать и такие химически устойчивые соединения, как углеводороды. Выявить способность микроорганизма окислять жидкие нелетучие углеводороды можно на плотной среде состава (г/л): KNO3 – 4,0; KH2PO4 – 0,6; Na2HPO4*12H2O – 1,4; MgSO4*7H2O – 0,8; выщелоченный агар – 2,0; pH 7,2. Среду стерилизуют в колбах при 1 атм. и разливают в чашки Петри толстым слоем. После того как среда застынет, в центре агаровой пластинки вырезают лунку. Для этой цели можно воспользоваться пробочным сверлом (диаметр 8–10 мм), которое предварительно стерилизуют в пламени горелки. Микромицеты высевают радиальными штрихами от лунки к периферии чашки. В лунку вносят 2–3 капли исследуемого углеводорода (стерилизуют фильтрованием). Чашки помещают в термостат строго горизонтально, не переворачивая. Через 7–10 суток отмечают наличие или отсутствие роста по штриху в сравнении с контролем – ростом на среде без углеводорода (Нетрусов, 2005).
2.5 Определение радиальной скорости роста
Определение радиальной скорости роста грибов проводили на плотной питательной среде за определенный промежуток времени. После 48 часов инкубации при 25±20С измеряют диаметр выросших на чашках колоний при помощи линейки. Эту операцию повторяют через каждые двое суток в течение двух недель.
За диаметр отдельной колонии в данный момент времени принимают среднее арифметическое измерение. Вычисление радиальной скорости проводят по формуле:
Kr = (r – ro) / (t – to),
где k радиальная скорость роста;
ro – радиус колоний в начальной момент времени to;
r – радиус колоний в момент времени t (Паников, 1991).
3. Исследование роста микромицетов рода Aspergillus на различных источниках углеродного питания
Объектами исследования явились различные виды микроскопических грибов рода Aspergillus: A. niger, A. ustus, A. terreus, A. flavus и A. fumigatus. В качестве источника углерода использовали из легкоусвояемых: сахара (сахароза, лактоза, ксилоза, арабиноза, галактоза, мальтоза), многоатомные спирты (глицерин, манит, сорбит), крахмал; из трудноразлагаемых (нефть, целлюлоза, гербицид, пестицид).
В результате посева исследуемых видов Aspergillus на среду Чапека с различными источниками углерода и среду Частухина с целлюлозой было установлено, что изменение экологических условий оказывает существенное влияние на развитие микромицетов. Была изучена способность штаммов использовать различные легкоусвояемые и трудноусвояемые источники углерода методом определения радиальной скорости роста выросших колоний. Оценка возможности потребления различных источников углерода показала, что они способны утилизировать многие источники углеродного питания, но ни один из исследуемых видов не разлагал такие трудноусвояемые вещества, как гербицид и пестицид. Динамика роста видов на разных средах при одинаковых условиях инкубации, не одинакова.
В таблице приложения и на рисунках 1–15 приведены зависимости радиальной скорости роста от времени у изученных грибов на различных источниках углерода.
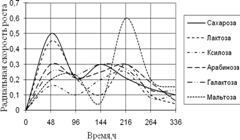
Рис. 1. Радиальная скорость роста A. niger на сахарах
Уже на первые сутки культивирования A. niger хорошо развивается во всех средах, кроме среды с ксилозой. Затем наблюдается скачок роста на средах с сахарозой и галактозой. К концу времени инкубации скорость роста колоний уменьшается и наибольшей становится на среде с мальтозой. Наибольшие биологические ритмы приходятся на 48–96 ч. и 144–264 ч. времени культивирования.
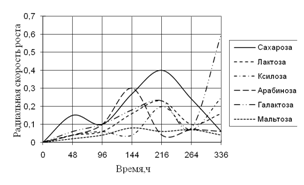
Рис. 2. Радиальная скорость A. terreus на сахарах
В первые дни культивирования наибольшая скорость роста A. terreus наблюдается на средах с сахарозой, арабинозой и галактозой. Затем происходит скачок роста на средах с сахарозой и арабинозой. На 14-е сутки культивирования наблюдается резкий скачок в росте на среде с галактозой. Биологические ритмы максимальны на 144–264 ч. дни инкубации.
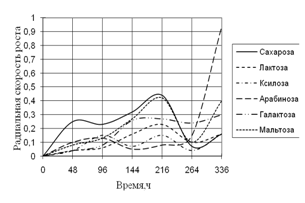
Рис. 3. Радиальная скорость роста A. fumigatus на сахарах
Наибольший рост A. fumigatus наблюдается на среде с сахарозой. Затем на этой среде рост уменьшается и происходит резкий скачок радиальной скорости на среде с арабинозой. К концу времени культивирования наблюдаются наибольшие биоритмы.
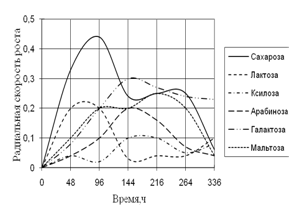
Рис. 4. Радиальная скорость роста A. flavus на сахарах
Из данных рисунка 4 видно, что A. flavus хорошо растет на среде с сахарозой. Но к концу культивирования наибольшая скорость роста наблюдается с галактозой. Максимальные биоритмы приходятся на начальный момент времени культивирования – 48–144 ч.
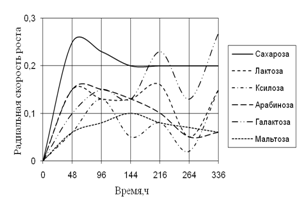
Рис. 5. Радиальная скорость роста A. ustus на различных сахарах
По данным рисунка 5 видно, что наибольшую скорость в течение всего времени инкубации A. ustus проявляет на среде с сахарозой, и резкий скачок в росте наблюдается на среде с галактозой. На протяжении всего времени инкубации рост A. flavus характеризуется высокими биоритмами.
Таким образом, сахара легко усваиваются всеми исследуемыми штаммами. Наибольшие биологические ритмы наблюдаются в течение всего времени экспозиции, как и в начальные часы, так и в конце инкубации.
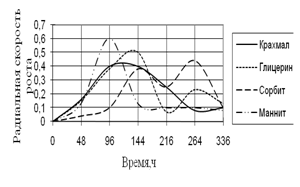
Рис. 6. Радиальная скорость роста A. niger
Наибольшая скорость роста A. niger наблюдается вначале культивирования на среде с маннитом. Затем скорость роста резко падает и становится минимальной на этой среде. На других источниках углерода также наблюдаются скачки роста, которые к концу инкубации заметно уменьшаются. Максимальные биоритмы приходятся на начальный момент времени культивирования – 48–144 ч.
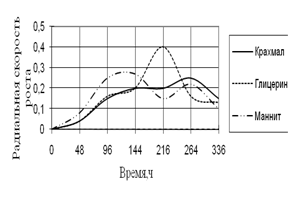
Рис. 7. Радиальная скорость роста A. terreus
Рост A. terreus наблюдается на всех легкоусвояемых источниках углерода, кроме среды с сорбитом. Вначале и в конце культивирования радиальная скорость роста примерно одинаковая. В середине инкубации наблюдаются скачки роста, особенно на среде с глицерином. Наибольшие биологические ритмы приходятся на 144–264 ч. времени культивирования.
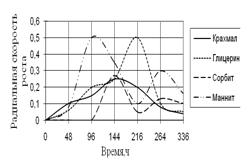
Рис. 8. Радиальная скорость роста A. fumigatus
По данным графика видно, что рост A. fumigatus имеет большие скачкообразные изменения, особенно проявляющиеся на средах с маннитом и глицерином. Наибольшая скорость роста проявляется на среде с маннитом, к середине инкубации наблюдается скачок роста на среде с глицерином. Биологические ритмы характеризуются большими колебаниями и приходятся на середину времени экспозиции – 96 ч. и 216 ч.
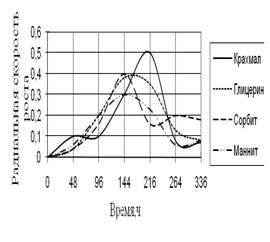
Рис. 9. Радиальная скорость роста A. flavus
Радиальная скорость роста A. flavus вначале и в конце культивирования примерно одинаковая. В середине инкубации наблюдаются скачки роста, особенно на среде с крахмалом. Высокие биоритмы наблюдаются на 144–264 ч. инкубации.
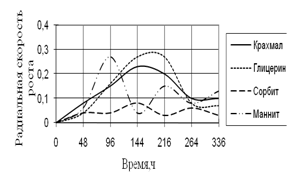
Рис. 10. Радиальная скорость роста A. ustus
Наибольшая скорость роста A. ustus наблюдается на средах с маннитом и глицерином. Скорость роста на среде с сорбитом наименьшая и в течение всего времени культивирования значительно не изменяется. Также как и A. fumigatus, A. ustus проявляет большие биоритмы на 144–216 ч.
В целом, рост микромицетов на многоатомных спиртах и крахмале характеризуется высокими биологическими ритмами, приходящимися на середину времени культивирования – 144–264 ч.
Ни один из исследуемых штаммов не растет на средах с единственными источниками углерода в виде пестицида и гербицида.
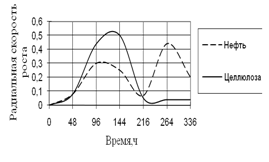
Рис. 11. Радиальная скорость A. niger на трудноразлагаемых источниках углерода
На трудноразлагаемых источниках углерода рост A. niger наблюдается на средах с целлюлозой и нефтью. В начале инкубации скорость роста на обеих средах одинакова, затем резко увеличивается на среде с целлюлозой, но к концу культивирования она становится минимальной, и происходит скачок роста на среде с нефтью. Максимальные биологические ритмы приходятся к середине времени культивирования – к 96–216 ч.
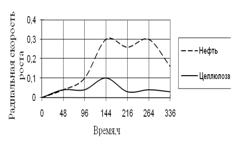
Рис. 12. Радиальная скорость роста A. terreus на трудноразлагаемых источниках углерода
Из трудноразлагаемых источников углерода A. terreus использует такие, как нефть и целлюлоза. Радиальная скорость роста на обоих источниках в начале культивирования была одинаковой, затем на среде с нефтью резко увеличивается к середине инкубации. На среде с целлюлозой резких скачков роста не наблюдается. Высокие биоритмы также находятся в центре экспозиции – 144–264 ч.
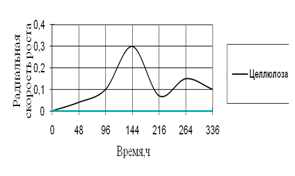
Рис. 13. Радиальная скорость роста A. fumigatus на трудноразлагаемых источниках углерода
На трудноусвояемых источниках углерода рост A. fumigatus наблюдается только на среде с целлюлозой. На 7-е сутки культивирования наблюдается резкий скачок роста, затем он уменьшается до начальной величины. Максимальные биоритмы приходятся на время инкубации – 96–216 ч.
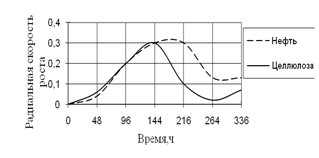
Рис. 14. Радиальная скорость A. flavus на трудноразлагаемых источниках углерода
Скорость роста на обоих источниках вначале культивирования была примерно одинаковой, максимальной – к середине инкубации, а затем она резко уменьшается. К концу культивирования наибольшая скорость роста A. flavus наблюдается на нефти.
Высокие биологические ритмы приходятся на середину времени экспозиции – 96 ч. и 216 ч.
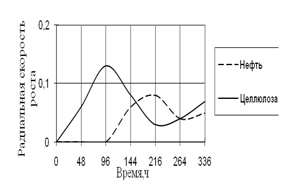
Рис. 15. Радиальная скорость роста A. ustus на трудноразлагаемых источниках углерода
Также как и другие виды, A. ustus проявляет свой рост только на средах с нефтью и целлюлозой. В начале культивирования на среде с целлюлозой наблюдается максимальная скорость роста. На 7-е сутки инкубации происходит скачок роста на среде с нефтью. К концу времени инкубации скорость роста A. ustus на обеих средах примерно одинаково. Наибольшие биоритмы характерны 48–144 ч.
Рост исследуемых штаммов на трудноразлагаемых источниках углерода характеризуются высокими биоритмами в середине экспозиции.
Таким образом, при определении способности штаммов использовать различные легкоусвояемые источники углерода, было выяснено, что на сахарах наибольшая скорость роста A. niger наблюдается на среде с мальтозой. A. terreus, A. fumigatus и A. ustus проявляют наибольшую скорость роста на средах с галактозой, а A. flavus – на среде с сахарозой. На крахмале и многоатомных спиртах все исследуемые штаммы проявляли высокую скорость роста, за исключением A. terreus, рост которого на среде с сорбитом вообще не наблюдался.
Из трудноразлагаемых источников углерода исследуемые штаммы утилизировали только нефть и целлюлозу, кроме A. fumigatus, который проявлял способность к росту только на среде с целлюлозой. Причем все виды имели высокую радиальную скорость роста на обеих средах.
Максимальные биологические ритмы при росте на сахарах наблюдаются в течение всего времени экспозиции. Это объясняется тем, что они легко усваиваются микромицетами. На крахмале, спиртах и трудноусвояемых источниках углерода наибольшие биоритмы приходятся на 96–216 ч. – середину инкубации.
Выводы
1) Была определена способность исследуемых штаммов использовать легкоразлагаемые источники углерода. При определении способности штаммов использовать различные легкоусвояемые источники углерода, было выяснено, что из сахаров A. niger отдает предпочтение мальтозе. A. terreus, A. fumigatus и A. ustus предпочитают среду с единственным источником углерода в виде галактозы, а A. flavus – среде с сахарозой. На крахмале и многоатомных спиртах все исследуемые штаммы проявляли высокую скорость роста, за исключением A. terreus, рост которого на среде с сорбитом вообще не наблюдался.
2) Определена способность штаммов использовать такие трудноразлагаемые источники углерода, как нефть и целлюлоза. Отсутствие роста на средах с гербицидом и пестицидом объясняется сильным токсическим действием их на микромицеты и не способностью изученных штаммов использовать данные вещества в качестве источника углерода.
СПИСОК ЛИТЕРАТУРЫ
1) Андреюк, Е.И. Микробная коррозия и ее возбудители [Текст] / Е.И. Андреюк, В.И. Билай, Э.З. Коваль, И.А. Козлова. – Киев: Наук. думка, 1980. – 286 с.; 22 см. – Библиогр.: с. 156. – 200 экз.
2) Бабьева, И.П. Изменения численности микроорганизмов в почвах при загрязнении тяжелыми металлами [Текст] / И.П. Бабьева, С.В. Левин, Н.С. Решетова // Тяжелые металлы в окружающей среде. – М.: Изд-во Моск. ун-та, 1980. – С. 115. – Библиогр.: с. 75.
3) Бабьева, Е.Н. Сравнительно-экологические исследования микромицетов из почв отдаленных географических районов [Текст] / Е.Н. Бабьева // Микология и фитопатология. Сер. 17. – 1983. – №2. – С. 452–453. Библиогр.: с. 452–453.
4) Биоповреждения [Текст] / Под ред. В.Д. Ильичева. – М.: Изд-во Моск. ун-та, 1987. – 352 с.; 24 см. – Библиогр.: с. 207–208. 200 экз. – ISBN 5–02634–675–3.
5) Блажеевская, Ю.В. Сравнительный анализ скорости радиального роста микромицетов, выделенных из различных экотопов [Текст] / Ю.В. Блажеевская, В.В. Вембер, Н.Н. Жданова // Микробиологический журнал. – 2002. Т. 64. – №3. – С. 3–11. – Библиогр.: с. 49–50.
6) Богомолова, Е.В. Морфологические особенности микроколониальных грибов, изолированных с поверхности камня [Текст] / Е.В. Богомолова, М.С. Зеленская, Д.Ю. Власов // Микология и фитопатология. Сер. 35. – 2001. – №3. – С. 6–13. – Библиогр.: с. 17.
7) Бухало, А.С. Высшие съедобные базидиомицеты в чистой культуре [Текст] / А.С. Бухало. – Киев: Наук. думка, 1988. – 144 с.; 21 см. Библиогр.: с. 76. – 3000 экз.
8) Великанов, Л.Л. Некоторые биохимические аспекты в экологии грибов [Текст] / Л.Л. Великанов, И.И. Сидорова // Успехи микробиологии. Сер. 3. – 1983. – №18. – С. 112–132. – Библиогр.: с. 128.
9) Воронин, Л.В. Микрофлора некоторых видов рыб Куйбышевского водохранилища [Текст] / Л.В. Воронин // Биология внутр. вод. – 1999 76. – С. 11–15. – Библиогр.: с. 13.
10)Гарибова, Л.В. Основы микологии: Морфология и систематика грибов и грибоподобных организмов [Текст]: учеб. пособие / Л.В. Гарибова, С.Н. Лекомцева М.: Товарищество научных изданий КМК, 2005. – 202 с.; 25 см. Библиогр.: с. 196–199. – 2000 экз. – ISBN 5–87317–265-X.
11)Григорьев, А.М. Изучение роста фрагментов мицелия Fusarium oxysporum в условиях разной кислотности среды [Текст] / А.М. Григорьев, М.В. Горленко, О.Е. Марфенина // Микология и фитопатология. 2004. – №3. – С. 29–35. – Библиогр.: с. 30–31.
12)Долгова, А.В. Рост колоний Penicillium chrysogenum Thom. при постоянных и переменных температурах [Текст] / А.В. Долгова, В.В. Зданович // Микология и фитопатология. Сер. 31. – 1997. – №1. – С. 52–56. – Библиогр.: с. 55.
13)Дудка, И.А. [Текст] Водные несовершенные грибы СССР / И.А. Дудка. Киев: Наук. думка, 1985. – 188 с.; 24 см. – Библиогр.: с. 154. 1500 экз. – ISBN 5–137–06374–4.
14)Евдокимова, Г.А. Микробиологическая активность почв при загрязнении тяжелыми металлами [Текст] / Г.А. Евдокимова // Почвоведение. – 1982. – №6. – С. 125 – 132. – Библиогр.: с. 130.
15)Звягинцев, Д.Г. Биология почв [Текст]: учебник / Д.Г. Звягинцев, И.П. Бабьева, Г.М. Зенова – 3-е изд., испр. и доп. – М.: Изд-во МГУ, 2005. – 445 с.: ил.; 25 см. – Библиогр.: с. 373–375 – 3000 экз. ISBN 5–211–04983–7.
16)Иванова, А.Е. Жизнеспособность фрагментов мицелия почвенных микроскопических грибов в разных экологических условиях [Текст]: автореф. канд. дис…; утверждена; защищена 30.03.99. / Иванова Анна Евгеньевна. – М.: МГУ, 1999. – 30 с.
17)Иванова, А.Е. Влияние экологических факторов на способность к росту фрагментов мицелия и прорастание спор микроскопических грибов [Текст] / А.Е. Иванова, О.Е. Марфенина // Микробиология. – 2001. – №2. – С. 235–240. Библиогр.: с. 236.
18)Коваль, Э.З. Микодеструкторы промышленных материалов [Текст] / Э.З. Коваль, Л.П. Сидоренко. – Киев: Наук. думка, 1989. – 192 с.; 22 см. – Библиогр.: с. 135–136. – 300 экз. – ISBN 5–015–02369–7.
19)Кочкина, Г.А. Радиальная скорость роста грибов в связи с их экологией [Текст] / Г.А. Кочкина, Т.Г. Мирчинк, П.А. Кожевин, Д.Г. Звягинцев // Микробиология. – 1978. – №5. – С. 964–965. – Библиогр.: с. 964.
20)Лилли, В. Физиология грибов [Текст] / В. Лилли, Г. Барнетт. Москва: Изд-во иностр. литературы, 1953. – 532 с.; 25 см. Библиогр.: с. 152, 174–184. – 2000 экз. – ISBN 5–248–00487–4.
21)Марьиновская, Ю.В. Микробиологическая деструкция целлюлозосодержащих отходов [Текст] / Ю.В. Марьиновская, Н.Н. Севастьянова // Микробиология. – 2006. – №3. – С. 75 – 81. – Библиогр.: с. 78.
22)Мирчинк, Т.Г. Почвенная микология [Текст]: учебник / Т.Г. Мирчинк: М.: Изд-во МГУ, 1988. – 220 с.; 22 см. – Библиогр.: с. 154 -165. – 2940 экз. – ISBN 5–211–00157–5.
23)Мюллер, Э. Микология [Текст] / Э. Мюллер, В. Леффлер; перевод с немецкого канд. биол. наук К.Л. Тарасова. – М.: Мир, 1993. – 535 с.; 25 см. – Библиогр.: с. 90–94, с. 139–140. – 2000 экз. – ISBN 5–214–01254–7.
24)Ниязова, Г.А. Концентрирование цинка и свинца различными микроорганизмами, обитающими в почве Сумсарского свинцово-цинкового субрегиона [Текст] / Г.А. Ниязова, С.В. Летунова, Б.Н. Золотарева // Микробиология. – 1982. – №4. – С. 650–656. – Библиогр.: с. 654.
25)Паников, Н.С. Кинетика роста микроорганизмов [Текст] / Н.С. Паников. М.: Наука, 1991. – 309 с.; 22 см. – Библиогр.: с. 245. – 1500 экз. – ISBN 3–271–00356–5.
26)Пидопличко, Н.М. Грибная флора грубых кормов [Текст] / Н.М. Пидопличко. Киев: Наук. думка, 1953. – 482 с.; 21 см. – Библиогр.: с. 246. 700 экз.
27)Практикум по микробиологии [Текст] / Под ред. А.И. Нетрусова. М.: Академия, 2005. – 608 с.; 28 см. – Библиогр.: с. 239–240. 5100 экз. – ISBN 5–7695–1809-X.
28)Романов, Ю.А. Биологические ритмы на разных уровнях биологической организации [Текст] / Ю.А. Романов // Проблемы космической биологии. – 1980. – №4. – С. 10–25. – Библиогр.: с. 11.
29)Сычугова, О.В. Рост и развитие микромицетов на сополимере этилена и винилацетата с добавками крахмала [Текст] / О.В. Сычугова, Н.Н. Колесникова // Вестн. Моск. ун-та. Сер. 16, Биология. – 2003. – №4. С. 27–31. Библиогр.: с. 28.
30)Терехова, В.А. [Текст] Микромицеты в экологической оценке водных и наземных экосистем / В.А. Терехова. – М.: Наука, 2007. – 215 с.; 21 см. Библиогр.: с. 33–35. – 2000 экз. – ISBN 5–453–06754–3.
31)Фомин, В.А. Биоразлагаемые полимеры: состояние и перспективы использования [Текст] / В.А. Фомин, В.В. Гузеев // Пластические массы. – 2001. – №2. – С. 42–46. – Библиогр.: с. 42–43.
32)Шаркова, Т.С. Цитохимическая характеристика спорулирующей и вегетативной зон культуры Trichothecium roseum F r. [Текст] / Т.С. Шаркова // Микология и фитопатология. – 1971. – №6. – С. 490–493. – Библиогр.: с. 492.
33)Шевцова, В.М. Программы развития и возможный принцип их генетического контроля у микромицетов рода Verticillium [Текст] / В.М. Шевцова // Микология и фитопатология. – 2001. – №21. – С. 73–81. – Библиогр.: с. 76.
34)Ellis, M.B. Dematiaceous hyphomycetes. Commonwealth. Kew [Text] / M.B. Ellis. – New York: Academic Press, 1971. – 608 p.; 25 cm. – Bib.: p. 246. – 3000 copy.
35)Lodder, J. The Yeasts, a taxonomical study. 2 ed. North – Holland [Text] / J. Lodder. – Amserdam, London: Publishing Corp, 1970. – 1385 p.; 21 cm. – Bib.: p. 642. – 2500 copy.
36)Barnett, J.A. A quide for identyifying and classifying yeasts [Text] / J.A. Barnett, R.W. Payne, D. Yarrow. – Cambridge, England: University Press, 1979. – 1315 p.; 26 cm. – Bib.: p. 756. – 1500 copy.
37)Hopper, H. Involment of clay type and pH in the mechanisms of soil suppressiveness to Fusarium wilt of flax [Text] / H. Hopper, C. Steinberg, C. Alabouvette // Soil Boil. Biochem. – 1995. – Vol. 27. – №7. – P. 955–967. – Bib.: p. 961.
38)Margollin, A.S. The effects of various carbohydrates upon the growth of some fungi, thesis [Text] / A.S. Margollin. – West Virginia University, 1942. – 327 p.; 25 cm. – Bib.: p. 234. – 5000 copy.
39)Moore, D. Metabolism and biochemistry of hyphal systems [Text] / D. Moore // Fungal morphogenesis. – 2001. – №4. – P. 26–134. – Bib.: p. 36.
40)Parton, W.J. Chemical aktivites of fungi [Text] / W.J. Parton, J.B. Stewart, C.V. Cole // Biogeochemistry. – 1988. – №5. – P. 109 – 131. – Bib.: p. 115.
41)Watanabe, T. Pictorial atlas of soil and seed fungi. Morphologies of cultured fungi end key to species [Text] / T. Watanabe. – Florida, 2000. – 411 p.
© 2009 База Рефератов