
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Контрольная работа: Синапсы. Центральная нервная система
Контрольная работа: Синапсы. Центральная нервная система
Физиология синаптической передачи
Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч. Шеррингтоном. Он, исследуя деятельность ЦНС, предположил еще в 1897 г., что нейроны между собой сообщаются с помощью специального механизма, который он и назвал синаптическим. Последующие годы подтвердили эту идею.
Классификация синапсов
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). Все синапсы ЦНС можно классифицировать следующим образом.
1. По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шенсрду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки).
2. По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
3. По конечному эффекту: тормозные и возбуждающие.
4. По механизму передачи сигнала: электрические, химические, смешанные.
5. Химические синапсы можно классифицировать:
а) по форме контакта – терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона);
б) по природе медиатора – холинергические (медиатор – ацетилхолин, АХ), адренергическис (медиатор – норадреналин, НА), дофаминергические (дофамин), ГАМК-ергические (медиатор – гаммааминомасляная кислота), глицинергические, глутаматергические, аспартатсргические, пептидергические (медиатор – пептиды, например, вещество Р), пуринергические (медиатор – АТФ).
Электрические синапсы. В настоящее время признают, что в ЦНС имеются электрические синапсы. С точки зрения морфологии электрический синапс представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно перескакивают через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД второй клетки. В целом, такие синапсы (они называются эфапсами) обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т. к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя заставить эффекторную клетку (клетку, которая управляется через данный синапс) тормозить свою активность. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Химические синапсы. По строению химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы), которая заполнена химическим веществом – медиатором. В синапсе различают пресинаптический элемент, который ограничен пресинаптической мембраной, постсинаптический элемент, который ограничен постсипаптической мембраной, а также внесинаптическую область и синаптическую щель, величина которой составляет в среднем 50 нм. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка – это синапс между нейронами, концевая пластинка – это постсинаптическая мембрана мионеврального синапса, моторная бляшка – это пресинаптичсское окончание аксона на мышечном волокне.
Общие принципы работы химического синапса
Любой химический синапс, независимо от природы медиатора и хеморецептора, активируется под влиянием потенциала действия, прибегающего к пресинапсу от тела нейрона. В результате – происходит деполяризация пресинаптической мембраны, что повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов кальция. В ответ на это происходит высвобождение квантов (выход из пресинапса) – 100–200 порций (квантов) медиатора. Выйдя в синаптическую щель, медиатор взаимодействует со специфическим рецептором постсинаптической мембраны, что вызывает изменение ионной проницаемости. В синапсах, в которых осуществляется возбуждение постсинантической структуры, обычно происходит повышение проницаемости для ионов натрия, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила определенное название: возбуждающий постсинаптический потенциал (ВПСП). Если его величина достаточно велика и достигает критического уровня деполяризации, то генерируется ПД. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и хлора). Это называется тормозным постсинаптическим потенциалом (ТПСГ1). В гиперполяризоваином состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или (если она обладала свойством автоматии) уменьшает спонтанную активность.
После каждого цикла проведения импульса медиатор разрушается, например, ацетилхолин разрушается ацстилхолинэстеразой, норадреналин разрушается моноаминоксидазой (МАО) или катсхол-0-метилтрансферазой (КОМТ), либо идет обратный захват в преси-наптическую структуру. В одних случаях захватывается неразрушенный медиатор (например, норадреналип), в других – его осколки (например, холин из ацетилхолина).
Синтез медиатора совершается в пресинаптическом элементе, куда попадают исходные продукты для синтеза и ферменты, необходимые в синтезе. Ферменты образуются в соме нейрона и по аксону, примерно со скоростью 6 мм/сутки, попадают в пресинапс, где используются в процессе синтеза медиатора. Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе и, следовательно, к снижению его функциональной способности.
Свойства химических синапсов
1. Односторонняя проводимость одно из важнейших свойств химического синапса. Асимметрия – морфологическая и функциональная – является предпосылкой для существования односторонней проводимости.
2. Наличие синаптической задержки: для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошло изменение постсинаптического потенциала (ВИСИ или ТПСП), требуется определенное время (синаптическая задержка). В среднем оно равно 0,2–0,5 мс.
3. Благодаря синаптическому процессу нервная клетка, управляющая данным постсинаптичсским элементом (эффектором), может оказывать возбуждающее воздействие или, наоборот, тормозное (это определяется конкретным синапсом).
4. В синапсах существует явление отрицательной обратной связи – антидромный эффект. Речь идет о том, что выделяемый в синаптическую щель медиатор может регулировать выделение следующей порции медиатора из этого же пресинаптического элемента путем воздействия на специфические рецепторы пресинаптичсской мембраны.
5. Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс. Если этот интервал до некоторых пор уменьшать (учащать подачу импульса по аксону), то на каждый последующий ПД ответ постсинаитической мембраны (величина ВПСП или ТПСП) будет возрастать (до некоторого предела). Это явление облегчает передачу в синапсе, усиливает ответ постсинаптического элемента (объекта управления) на очередной раздражитель; оно получило название «облегчение» или «потенциация». В основе его лежит накопление кальция внутри пресинапса. Если частота следования сигнала через синапс очень большая, то из-за того, что медиатор не успевает разрушиться или удалиться из синаптической щели, возникает стойкая деполяризация или катодическая депрессия – снижение эффективности синаптической передачи. Это явление называется депрессией. Если через синапс проходит много импульсов, то в конечном итоге постсинаптическая мембрана может уменьшить ответ на выделение очередной порции медиатора. Это называется явлением дссепситизации утратой чувствительности. В определенной мере десеиситизация похожа па процесс рефрактерности (утрата возбудимости). Синапсы подвержены процессу утомления. Возможно, что в основе утомления (временного падения функциональных возможностей синапса) лежат: а) истощение запасов медиатора, б) затруднение выделения медиатора, в) явление дссенситизации. Т. о., утомление – это интегральный показатель.
В мозге имеется ряд медиаторов, вызывающих возбуждение нейрона: норадреналин (его продуцируют адренергические нейроны), дофамин (дофаминергические нейроны), серото-нин, пептиды (пептидергические), глутаминовая кислота, аспарагиновая кислота и т.д. Во всех этих случаях выделяющийся медиатор взаимодействует со специфическим рецептором, в результате чего меняется проницаемость для ионов натрия, калия или хлора, и в итоге развивается деполяризация (ВПСП). Если она достигает критического уровня деполяризации, то возникает ПД (возбуждение нейрона).
Тормозные синапсы образованы специальными тормозными нейронами (точнее, их аксонами). Медиатором могут быть глицин, гамма-аминомасляная кислота (ГАМК) и ряд других веществ. Обычно глицин вырабатывается в синапсах, с помощью которых осуществляется постсинаптическое торможение. При взаимодействии глицина как медиатора с глициновыми рецепторами нейрона возникает гиперполяризация нейрона (ТПСП) и, как следствие, – снижение возбудимости нейрона вплоть до полной его рефрактсрности. В результате этого возбуждающие воздействия, оказываемые через другие аксоны, становятся малоэффективными или неэффективными. Нейрон выключается из работы полностью.
Центральная нервная система
Центральная нервная система (ЦНС) – это совокупность нервных образований спинного и головного мозга, обеспечивающих восприятие, обработку, передачу, хранение и воспроизведение информации с целью адекватного взаимодействия организма и изменений окружающей среды, организации оптимального функционирования органов, их систем и организма в целом.
Центральная нервная система человека представлена спинным продолговатым, средним, промежуточным мозгом, мозжечком, базальными ганглиями и корой головного мозга.
Нейрон и нейроглия
Нейрон является функциональной единицей мозга. Наряду с нейронами в мозге имеются клетки глии – олигодендроциты и астроциты.
Нейрон имеет дендриты, аксон, сому. Для физиолога очень важно понятие об аксонном холмике и начальном сегменте. Именно в этом месте происходит возбуждение нейрона, так как эта часть обладает наибольшей возбудимостью.
Нейрон имеет множество синапсов, через которые он получает возбуждение и тормозные воздействия от других нейронов. Благодаря этому нейрон может получать в больших количествах информацию.
Нейрон может находиться в различных состояниях: а) в состоянии покоя – практически отсутствуют колебания мембранного потенциала, ПД не генерируется; б) в состоянии активности – генерировать потенциалы действия (для нейронов характерна генерация серии или пачки импульсов). Состояние активности может быть индуцировано за счет поступления к нейрону импульсов от других нейронов или быть спонтанным (автоматия). В этом случае нейрон играет роль пейсмекера (водителя ритма). Такие нейроны имеются в ряде центров, например, в центре дыхания; в) в состоянии торможения – оно проявляется в том, что нейрон прекращает свою импульсную активность (нейрон – пейсмекер, или нейрон, получающий возбуждающие воздействия). В основе торможения лежит явление гиперполяризации нейрона (это характерно для постсинаптического торможения).
|
|
Возможные состояния нейрона1 – поляризация, 2 – гиперполяризация, 3 – деполяризация; МП – мембранный потенциал, ТПСП – тормозящий постсинаптический потенциал, ПД потенциал действия |
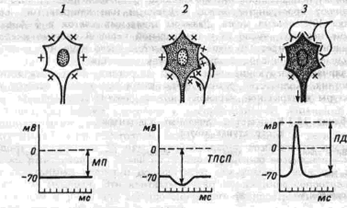
В нервной клетке при развитии потенциала действия возникает период рефрактерности или невозбудимости. Это явление заключается в том, что на фоне потенциала действия и еще некоторое время после его завершения, нельзя вызвать второй разряд клетки, невзирая на силу применяемого раздражения. Время, в течение которого нервная клетка невозбудима, называется абсолютной рефрактерностью. Затем клетка приобретает возможность реагировать на прилагаемые раздражения, но впервые возникающий после периода абсолютной рефрактерности потенциал имеет меньшую амплитуду, так как возникает при меньшем уровне заряда мембраны. Периоды появления неполных ответов называются периодами относительной рефрактерности.
Функционально нейроны делят на три типа: афферентные, промежуточные и эфферентные. Первые – выполняют функцию получения и передачи информации в вышележащие структуры ЦНС, вторые – обеспечивают взаимодействие между нейронами одной структуры, третьи – за счет длинного аксона передают информацию в нижележащие структуры ЦНС, в нервные узлы, лежащие за ее пределами, и в органы организма.
По форме нейроны делят на моно-, би- и мультиполярные. По химической характеристике выделяемых в окончаниях аксонов веществ, отличают нейроны: холинэргические, пептидэргические, норадреналинэргические, дофаминэргические, серотонинэргические и др.
Важной характеристикой нейронов является их чувствительность к разным раздражителям. По этому признаку нейроны делят на моно-, би- и полисенсорные.
Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей модальности. Например, значительная часть нейронов первичной зрительной коры реагирует только на световое раздражение сетчатки глаза.
Бисенсорные нейроны располагаются преимущественно во вторичных зонах коры анализатора и могут реагировать как на сигналы своей, так и на сигналы другой модальности. Например, нейроны вторичной зрительной коры реагируют на зрительные и слуховые раздражения. Полисенсорные нейроны – это чаще всего нейроны ассоциативных зон мозга. Они способны реагировать на раздражение слуховой, зрительной, кожной и др. анализаторных систем.
Функции нейроглии. Глия – структура нервной системы, образованная специализированными клетками различной формы, которые заполняют пространства между нейронами или капиллярами, составляя 10% объема мозга. Размеры глиальных клеток в 3–4 раза меньше нервных, число их в центральной нервной системе млекопитающих достигает 140 млрд. С возрастом число нейронов в мозгу уменьшается, а число глиальных клеток увеличивается.
Различают следующие виды глии: астроглия, олигодендроглия, микроглия.
Астроглия представлена многоотростчатыми клетками. Их размеры колеблются от 7 до 25 мкм. Большая часть отростков заканчивается на стенках сосудов. Астроглия служит опорой нейронов, обеспечивает репаративные процессы нервных стволов, изолирует нервное волокно, участвует в метаболизме нейронов.
Олигодендроглия это клетки, имеющие один отросток. Количество олигодендроглии возрастает в коре от верхних слоев к нижним. В подкорковых структурах, в стволе мозга олигодендроглии больше, чем в коре. Она участвует в миелинизации аксонов, в метаболизме нейронов.
Микроглия самые мелкие клетки глии, относятся к блуждающим клеткам. Они образуются из структур оболочек мозга, проникают в белое, а затем и в серое вещество мозга. Микроглиальные клетки способны к фагоцитозу.
Одной из особенностей глиальных клеток является их способность к изменению своего размера. Изменение размера глиальных клеток носит ритмический характер: фазы сокращения – 90 с, расслабления – 240 с, т.е. это очень медленный процесс. Средняя частота ритмических изменений варьирует от 2 до 20 в час. При этом отростки клетки набухают, но не укорачиваются в длине.
Физиологические процессы в нервной системе во многом зависят от миелинизации волокон нервных клеток. В центральной нервной системе миелинизация обеспечивается олигодендроглией, а в периферической – шванновскими клетками.
Торможение в ЦНС его виды и предполагаемые механизмы
Впервые идею о том, что в ЦНС помимо процессов возбуждения существует процесс торможения, высказал И.М. Сеченов.
Если рассмотреть «архитектуру» использования тормозных нейронов при организации нейронных сетей, цепей и рефлекторных дуг, то можно выделить ряд вариантов этой организации (это отражается в названии данного вида торможения).
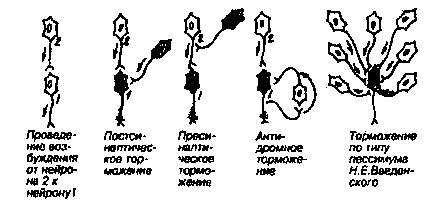
1. Реципрокное торможение. Как пример: сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на альфа-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона разгибателя. Явление открыто Ч. Шеррингтоном.
2. Возвратное торможение. Альфа-мотонейрон, к примеру, посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС – она заканчивается на тормозном нейроне (клетка Реншоу) и активирует ее. Тормозной нейрон вызывает торможение альфа-мотонейрона, который запустил всю эту цепочку. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя тормозит.
3. Существует ряд вариантов возвратного торможения, в частности, латеральное торможение. Суть его сводится к тому, что активируется, например, фоторецептор, он активизирует биполярную клетку, и одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглиозной клетке. Этим самым происходит «вытормаживание» информации в соседних участках. Таким способом создаются условия для четкого видения предмета (две точки на сетчатке рассматриваются как две раздельные точки в том случае, если между ними есть невозбужденные участки).
Особенности распространения возбуждения в ЦНС
Рассматривая основные принципы и особенности распространения возбуждения в ЦНС, а по сути, и свойства нервных центров, можно выделить следующие моменты.
1. Одностороннее проведение возбуждения. В ЦНС – в ее центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении, например, от афферентного нейрона к эфферентному, а не наоборот. Обусловлено это особенностями расположения и характером функционирования химического синапса.
2. Суммация возбуждений (аналогично можно говорить и о суммации торможения). На нейроне в области его аксонального холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона. Если с определенным интервалом к нейрону в точку А приходят импульсы, они вызывают генерацию в этой области ВПСП. Если эти ВПСП не достигают критического уровня деполяризации, то потенциал действия не возникает. Если же частота следования достаточно большая, то происходит в этом месте суммация ВПСП, при достижении ВПСП критического уровня деполяризации возникает ПД, нейрон возбуждается. Это явление носит название временной суммации (происходит суммация следов возбуждения во времени). В ЦНС имеет место и пространственная суммация: возбуждения, приходящие в точку В, А, С нейрона (даже если они сами по себе – подпороговые), при одновременном появлении у данного нейрона могут привести к его возбуждению при условии, что суммированный ВПСП достигает или превышает критический уровень деполяризации.
3. Явление окклюзии: один и тот же нейрон может передавать сигналы на ряд других нейронов, в результате чего возникает определенный эффект (например, происходит активация 10 мышечных волокон в мышце, вследствие чего мышца развивает напряжение, равное 100 мгс). Второй нейрон возбуждает тоже 10 других волокон (100 мгс). Но если оба нейрона возбуждать одновременно, то суммарная активность мышцы будет 180 мгс. Почему? Оказывается, часть волокон у них были общими (т. с. нейрон 1 и 2 передавали информацию на одни и те же волокна). Это явление получило название окклюзии или закупорки.
4. Трансформация ритма возбуждения. В отличие от скелетной мышцы или аксона нейрон способен Tpai формировать ритм возбуждений, приходящих к нему. Например, поступает импульс, идущий с частотой 25 Гц, а нейрон в ответ на это, возбуждаясь, генерирует 50 имп/сек (50 Гц), или наоборот, поступает 100 имп/сек, а выходят 40 имп/сек.
5. Последействие: один из вариантов этого свойства – длительное циркулирование импульсов но «нейронной ловушке». Итальянский физиолог Лоренто де Но обнаружил это явление: поступивший импульс может минутами или часами пробегать небольшой отрезок нейронной цепи. Благодаря этому, как полагают некоторые авторы, происходит перевод следа (энграммы) из краткосрочной памяти в долгосрочную.
6. Утомление нервных центров: это одно из важных свойств ЦНС. Оно обусловлено особенностями синаптической передачи в ЦНС: при длительном возбуждении одного и того же нейрона в синапсе может снизиться содержание медиатора, что приведет к снижению работоспособности нейрона.
Принципы координационной деятельности ЦНС
Координация это объединение действий в единое целое, объединение различных нейронов в единый функциональный ансамбль, решающий конкретную задачу. Координация способствует реализации всех функций ЦНС. Выделяют следующие принципы координации (их много, в лекции даются наиболее важные).
1. Явление конвергенции (концентрации) или принцип общего конечного пути. Многие нейроны оказывают свое воздействие на один и тот же нейрон, т. с. имеет место схождение потоков импульсов к одному и тому же нейрону. Ч. Шеррингтон называл это «принцип общего конечного пути». Например, сокращение мышцы (за счет возбуждения альфа-мотонейрона) можно вызвать путем растяжения этой мышцы (рефлекс мышечных веретен) или путем раздражения кожных рецепторов (сгибательный рефлекс) и т.п.
2. Явление дивергенции (иррадиации). Каждый нейрон за счет вставочных нейронов и многочисленных ветвлений (дивергенции) дает поток импульсов ко многим нейронам. Если бы этот механизм не ограничивался торможением, то благодаря дивергенции отсутствовала бы возможность координации работы ЦНС. Но тормозные процессы ограничивают дивергенцию и делают процессы управления точными. Когда торможение снимается, то имеет место полная дискоординация в деятельности ЦНС (например, при столбняке).
3. Принцип рсципрокной иннервации (см. Торможение; реципрокное торможение).
4. Принцип обратной связи и копий эфферентаций. Это один из важнейших принципов координации: невозможно точно координировать, управлять, если отсутствует обратная связь, т.е. данные о результатах управления. Осуществляется эта связь за счет потока импульсов с рецепторов. Этот принцип широко обсуждается в физиологии ЦНС, о нем уже говорил И.М. Сеченов, много внимания ему уделил П.К. Анохин.
Копия эфферентации. Для управления важно иметь информацию о том, какие команды посылаются на периферию. Известно, что в системах, управляющих скелетными мышцами, каждый отдел, посылая сигнал управления к работающей мышце, одновременно сообщает об этом вышележащему отделу. Это вариант обратной связи.
5. Принцип доминанты. Был открыт А.А. Ухтомским. Речь идет о том, что среди рефлекторных актов, которые могут быть выполнены в данный момент времени, имеются рефлексы, реализация которых представляет наибольший «интерес» для организма, т.е. они в данный момент времени самые важные. Поэтому эти рефлексы реализуются, а другие – менее важные – тормозятся.
6. Пластичность нервных центров. Для врача особенно важно знание этого свойства (принципа): при повреждении отдельных центров мозга их функция может перейти к другим структурам мозга (конечно, если повреждение центра не связано с наступлением смерти, что, например, бывает при нарушении дыхательного центра). Замещение утраченной функции – важнейшее приобретение ЦНС (известно, что нейроны ЦНС, как правило, не восстанавливаются) – оно позволяет восстанавливать утраченные свойства. Показано, что процесс возмещения утраченных функций осуществляется при обязательном участии коры больших полушарий: у животных, которым после восстановления нарушенных функций удаляли кору, вновь имела место утрата этой функции.
7. Принцип субординации или соподчинения. В ЦНС имеют место иерархические взаимоотношения начальник (кора) и подчиненные (сверху вниз – базальные ганглии, средний мозг, продолговатый, спинной) и соподчинение – нижележащий отдел подчиняется указаниям вышележащего отдела.
Информационная функция нервной системы. Процесс восприятия сигнала, обработка его и посылка к исполнительной системе связаны с кодированием, «шифрованием» информации. Это происходит уже на уровне отдельной нервной клетки.
Для процесса передачи информации большое значение имеет скорость ее передачи. Важное значение в передаче информации имеет также надежность. Для обеспечения надежности передачи информации в нервной системе используется принцип структурной и функциональной избыточности. Структурная избыточность выражается в дублировании каналов связи, дублировании элементами, реагирующими на данный сигнал, дублировании системами, включающимися в реакцию. Функциональная избыточность обеспечивается «излишним» числом импульсов в разряде нервной клетки, существенным повышением возбудимости нервных структур и др.
Считают, что нервные импульсы передают информацию двоичным кодом (наличие импульса отсутствие импульса). Из такого представления следует, что количество информации определяется числом импульсов в единицу времени.
© 2009 База Рефератов