
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Курсовая работа: Сахарный диабет. Нарушение обмена белков, углеводов и липидов при сахарном диабете
Курсовая работа: Сахарный диабет. Нарушение обмена белков, углеводов и липидов при сахарном диабете
ПЕНЗЕНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ПЕНЗЕНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ ИНСТИТУТ
Курсовая работа по биологической химии на тему:
САХАРНЫЙ ДИАБЕТ. НАРУШЕНИЕ ОБМЕНА БЕЛКОВ, УГЛЕВОДОВ И ЛИПИДОВ ПРИ САХАРНОМ ДИАБЕТЕ
Пенза 2004
СОДЕРЖАНИЕ
Введение
Инсулин и глюкагон как регуляторы депонирования и мобилизации углеводов и жиров
Секреция инсулина
Нарушения метаболизма углеводов и липидов при сахарном диабете
Коматозные состояния как результат нарушения обмена углеводов и жиров при сахарном диабете
Гликозилирование белков при сахарном диабете
Нарушение белкового обмена
Список литературы
ВВЕДЕНИЕ
Сахарный диабет является следствием нарушения инсулиновой регуляции функций ряда клеток организма. Поздние осложнения диабета: микроангиопатии (нефропатия, ретинопатия и др.) и макроангиопатии — часто приводят к ранней инвалидизации. Сахарный диабет — распространенная болезнь, занимает третье место среди причин смертности после сердечно-сосудистых заболеваний и рака. В мире около 100 млн. человек больны сахарным диабетом; каждые 10—15 лет число больных диабетом во всех странах мира удваивается. Наибольшему риску заболеть сахарным диабетом подвержены население развивающихся стран и группы малообеспеченных лиц в индустриально развитых странах. Диабетом II типа заболевают в зрелом возрасте, обычно после 40 лет. Он развивается постепенно, симптомы выражены умеренно, острые осложнения редки. Диабет I типа начинается обычно в юношеском возрасте, иногда в детстве, редко у взрослых. Протекает гораздо тяжелее, чем диабет II типа. При недостаточном врачебном контроле нередко развиваются острые осложнения. Распространенность диабета I типа почти в 10 раз меньше, чем диабета II типа. Сахарный диабет вследствие высокой распространенности, ранней инвалидизации и уменьшения продолжительности жизни больных является одной из важнейших медико-социальных проблем. Изучение механизмов инсулиновой регуляции, этиологии и патогенеза сахарного диабета, поиски новых методов лечения проводятся в мире очень широко и интенсивно. В последнее время главные задачи исследований — переход от диагностики диабета к его предсказанию, от лечения к предупреждению.
ИНСУЛИН И ГЛЮКАГОН КАК РЕГУЛЯТОРЫ ДЕПОНИРОВАНИЯ И МОБИЛИЗАЦИИ ГЛИКОГЕНА И ЖИРОВ
Инсулин участвует в регуляции таких клеточных процессов, как метаболизм, трансмембранный перенос ионов, аминокислот, глюкозы, синтез и распад белков. С влиянием на ядерные процессы — репликацию и транскрипцию – связано участие инсулина в регуляции клеточной пролиферации и дифференцировки, а также трансформации клеток. В патогенезе основных клинических проявлений сахарного диабета в наибольшей мере проявляется нарушение инсулиновой регуляции обмена глюкозы, жиров и аминокислот, связанного с энергетическим обменом. В результате согласованной работы разных органов и систем в организме поддерживается энергетический гомеостаз, под которым понимают соответствие между потребностью в энергии и обеспеченностью организма энергоносителями. Гомеостаз сохраняется даже при существенных изменениях в приеме пищи и энергетических затратах. Инсулин, а также тесно взаимодействующий с ним «контринсулярный» гормон глюкагон — главные регуляторы изменений метаболизма при смене состояний пищеварения и голодания (абсорбтивное и постабсорбтивное состояния). На пищеварение приходится 10—15 ч в сутки, а расход энергии происходит в течение всех 24 ч (с определенным снижением в часы ночного сна). Поэтому часть энергоносителей во время пищеварения складируется для использования в постабсорбтивном состоянии. Печень, жировая ткань и мышцы — главные органы, связанные с этими изменениями. Режим запасания включается после приема пищи и сменяется режимом мобилизации запасов после завершения пищеварения. Следовательно, у человека при обычном трехразовом питании смена режимов происходит трижды за сутки. Однако смена режимов выражена нечетко, поскольку в течение дня промежутки между приемами пищи небольшие (5 – 6 ч) и постабсорбтивный период едва успевает начаться (если вообще успевает), как наступает время очередного приема пищи. Типичным постабсорбтивным состоянием считают состояние утром до завтрака, после примерно десятичасового ночного перерыва в приеме пищи. Еще более наглядна модель ритма питания, которой придерживался великий немецкий философ Э. Кант: он принимал пищу один раз в сутки. За сутки исчерпываются запасы гликогена в организме, единственным источником глюкозы становится глюконеогенез, глюкоза используется преимущественно нервными клетками, в то время как почти все другие клетки получают энергию за счет окисления жирных кислот, а также кетоновых тел, образующихся в печени из жирных кислот. Такое состояние можно считать как постабсорбтивное или как кратковременное голодание. Эту модель (рис.1) мы и будем иметь в виду, рассматривая смену режимов обмена энергоносителей.
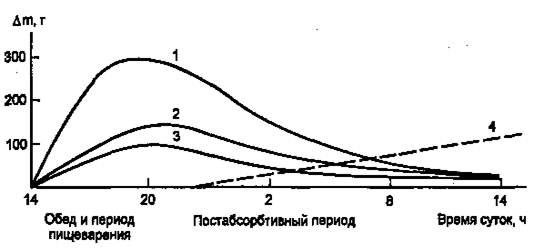
Рисунок 1. Изменение количества энергоносителей в организме человека (в тканях, не в желудке и кишечнике) в течение суток после однократного приема пищи. (1 — гликоген; 2 — жиры; 3 — аминокислоты/белки; 4 — изменение скорости глюконеогенеза, г/сут.)
Мышечная работа во время пищеварения замедляет процессы запасания, так как в мышцах непосредственно расходуется часть поступающих из кишечника продуктов переваривания. В постабсорбтивном состоянии мышечная работа стимулирует мобилизацию запасов, главным образом жиров. В регуляции изменений, связанных со сменой покоя и мышечной работы, важная роль принадлежит адреналину.
Потребление глюкозы клетками происходит при участии специальных белков-переносчиков (их называют также рецепторами глюкозы), образующих гидрофильные трансмембранные каналы. Существует два основных механизма переноса глюкозы: активный транспорт, зависящий от градиента концентраций ионов Na+, и облегченная диффузия. Соответственно есть два основных типа рецепторов глюкозы. Рецепторы, зависимые от концентрации ионов Na+, обнаруживаются только в почках и кишечнике, они обеспечивают реабсорбцию глюкозы из почечных канальцев и всасывание ее из просвета кишечника против градиента концентрации. Рецепторы облегченной диффузии (глюкозные транспортеры — ГЛЮТ) есть во всех тканях.
В тканях человека обнаружено пять разных ГЛЮТ:
ГЛЮТ-1 — в плаценте, мозге, почках, толстой кишке, в Р-клетках островков Лангерганса; меньше их в жировой ткани и мышцах;
ГЛЮТ-2 — преимущественно в печени, энтероцитах, в проксимальных тубулярных клетках почек (все эти клетки выделяют глюкозу в кровь); в Р-клетках панкреатических островков (островков Лангерганса). Возможно, участвует в стимуляции глюкозой секреции инсулина;
ГЛЮТ-3 во многих тканях, включая мозг, плаценту, почки;
ГЛЮТ-4 — единственный переносчик, регулируемый инсулином; содержится только в мышцах (скелетных и сердечной) и жировой ткани (инсулинзависимые ткани);
Все рецепторы могут находиться как в плазматической мембране клетки, так и в мембранных везикулах в цитоплазме. Количество рецепторов 1, 2, 3 и 5 в плазматической мембране изменяется в узких пределах и не зависит от концентрации инсулина. Напротив, ГЛЮТ-4 (и в гораздо меньшей степени ГЛЮТ-1) в отсутствие инсулина практически полностью находятся в цитозольных везикулах. Стимуляция клеток инсулином приводит к транслокации везикул к плазматической мембране и их слиянию, в результате чего рецепторы оказываются встроенными в плазматическую мембрану. Как показано в экспериментах с жировыми и мышечными клетками, скорость потребления глюкозы при этом увеличивается в 30—40 раз. При снижении концентрации инсулина в среде рецепторы вновь возвращаются в цитозоль.
Основными энергоносителями являются глюкоза и жирные кислоты. На рис. 2 представлены пути превращений глюкозы и жиров, а также белков и аминокислот.
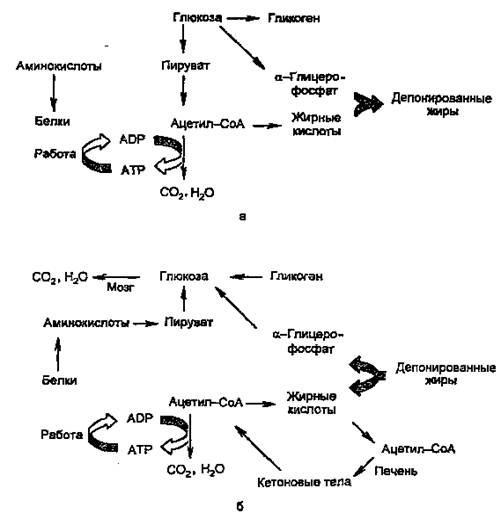
Рисунок 2. Изменение метаболизма основных энергоносителей при смене абсорбтивного (а) и постабсорбтивного (б) состояний.
Как видно из рисунка, при смене режимов многие процессы меняют направление на противоположное. За каждой из стрелок — серия реакций; ферменты, катализирующие ключевые реакции (лимитирующие скорость данной метаболической цепи), находятся под контролем многих регулирующих механизмов, включающих в качестве первого (внеклеточного) вестника сигнала главным образом инсулин и глюкагон, а также адреналин и кортизол.
Первичными сигналами для смены состояний являются изменение концентрации глюкозы в крови и вызванные этим реципрокные изменения концентраций инсулина и глюкагона. Регуляцию метаболизма инсулином и глюкагоном невозможно рассматривать по отдельности. В крови постоянно присутствуют оба гормона, однако изменяются их относительные концентрации. Действие каждого из них часто направлено на одни и те же конкретные мишени. Например, инсулин через путь Ras одновременно активирует гликогенсинтазу и ингибирует гликогенфосфорилазу, а глюкагон через сАМР-зависимые протеинкиназы одновременно ингибирует гликогенсинтазу и активирует гликогенфосфорилазу. Другой пример: инсулин сокращает не базальную скорость глюконеогенеза, а только скорость, стимулированную глюкагоном. На рис. 3 показаны некоторые другие мишени метаболических путей глюкозы в печени, общие для инсулина и глюкагона. Кроме того, инсулин снижает секрецию и самого глюкагона.
Глюкоза проникает в гепатоциты путем облегченной диффузии при участии ГЛЮТ-2, не зависимого от инсулина и имеющего высокую Км. В гепатоцитах глюкоза быстро превращается в глюкозо-6-фосфат глюкокиназой (гексокиназой IV), которая тоже имеет высокую Км (12 мМ) и не ингибируется продуктом реакции (в отличие от гексокиназ I, II и III). Далее глюкозо-6-фосфат может использоваться по трем направлениям: синтез гликогена, гликолиз, пентозофосфатный путь. Следует отметить, что ацетил-СоА, образующийся из глюкозы, используется для синтеза жирных кислот и жиров. Все эти пути стимулируются инсулином на пре- или посттрансляционном уровне.
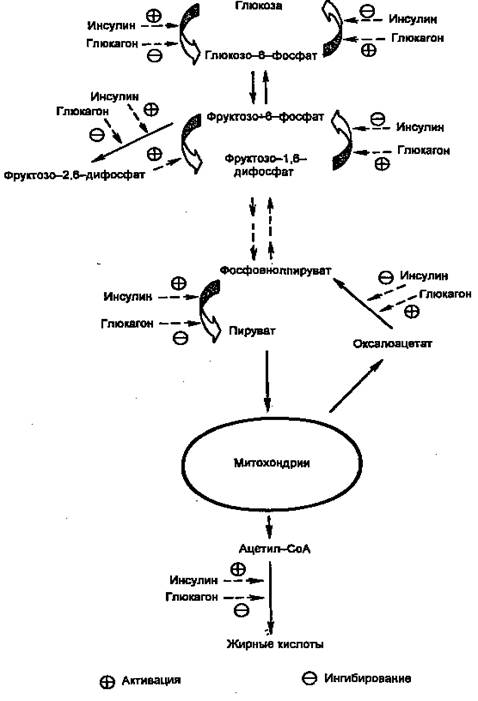
Рисунок 3. Действие инсулина и глюкагона на метаболизм глюкозы в печени.
Регуляция на претрансляционном уровне в свою очередь может быть двух типов: стимуляция транскрипции и повышение стабильности мРНК. В печени необратимые реакции гликолиза, а также синтез гликогена и синтез жиров стимулируются инсулином и подавляются глюкагоном. Наоборот, необратимые стадии глюконеогенеза подавляются инсулином и стимулируются глюкагоном. Подобная ситуация имеет место и в метаболизме жиров и аминокислот (белков): инсулин стимулирует их синтез, а глюкагон — мобилизацию. Поэтому направление метаболических процессов в сторону запасания или мобилизации зависит не столько от абсолютной концентрации гормона, сколько от отношения их концентраций ([инсулин]/[глюкагон], инсулин/глюкагоновый индекс). Для того, чтобы понять механизмы метаболических нарушений при инсулин-зависимом диабете, необходимо рассмотреть механизмы синтеза и секреции инсулина, так как нарушение этих процессов одна из причин развития заболевания.
СИНТЕЗ И СЕКРЕЦИЯ ИНСУЛИНА
Молекула инсулина построена из двух пептидных цепей: цепь А содержит 21 аминокислотный остаток, цепь Б — 30 остатков. Цепи соединены между собой двумя дисульфидными мостиками (рис. 7.6). Инсулины многих животных очень сходны по первичной структуре. С инсулином человека наиболее сходен инсулин свиньи, различие в одной позиции: в цепи Б в 30-й позиции (С-концевой остаток), у человека Тре, у свиньи — Ала.
Инсулин образуется из препроинсулина в результате посттрансляционной модификации. Ген препроинсулина в геноме человека представлен единственной копией. В настоящее время интенсивно изучаются строение промоторной области и механизмы регуляции гена инсулина.
Синтез препроинсулина происходит на полирибосомах, связанных с эндоплазматическим ретикулумом. Препроинсулин проникает в люмен ретикулума, где от него отщепляется лидирующая последовательность — N-концевой фрагмент, содержащий 24 аминокислотных остатка. Образовавшийся проинсулин (86 аминокислотных остатков) перемещается затем в аппарат Гольджи, где упаковывается в секреторные гранулы. В аппарате Гольджи и секреторных гранулах происходит превращение проинсулина в инсулин. В этом превращении участвуют две эндопептидазы: прогормонконвертазы 2 и 3 (ПГ2 и ПГЗ; последнюю называют также ПГ1). Эти ферменты расщепляют связи Арг32—Глу33 и Арг65—Гли66. Затем С-концевые остатки Apr и Лиз отщепляются карбоксипептидазой Е (КП-Е; известна также как КП-Н). Этот фермент есть во многих других органах, участвует в процессинге ряда гормонов и нейромедиаторов.
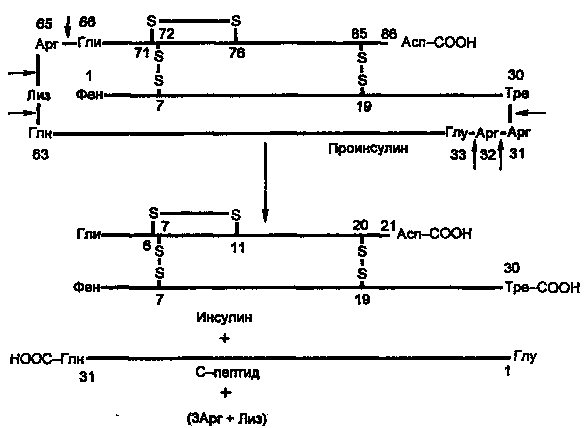 Рисунок
4. Процессинг проинсулина.
Рисунок
4. Процессинг проинсулина.
Таким образом, в секреторных гранулах содержатся (и секретируются из них) инсулин и С-пептид в эквимолярных количествах. Долгое время С-пептид рассматривали как физиологически неактивное вещество. Недавно было обнаружено, что в физиологических концентрациях он стимулирует потребление глюкозы клетками мышц здорового человека и больных ИЗСД примерно в такой же мере, как инсулин.
Глюкоза регулирует экспрессию гена инсулина, а также генов других белков, участвующих в обмене основных энергоносителей. Транскрипция ряда генов, связанных с метаболизмом, активируется в поджелудочной железе, печени и жировых клетках при потреблении пищи, содержащей углеводы.
Действие глюкозы может быть прямым, когда сама глюкоза или ее метаболиты непосредственно взаимодействуют с аппаратом регуляции гена, или вторичным, обусловленным влиянием глюкозы на секрецию гормонов, главным образом инсулина и глюкагона. Однако выяснить, что является регулятором — инсулин или глюкоза, можно только при использовании клеточных культур, позволяющих строго контролировать содержание этих веществ в среде (см. табл.).
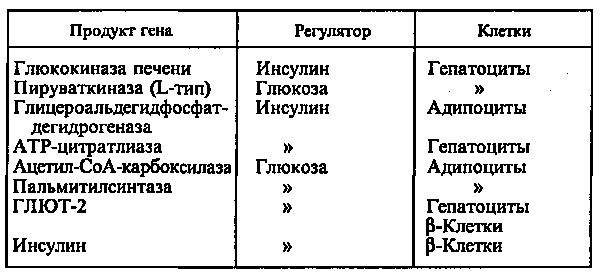
При стимуляции глюкозой инсулин быстро освобождается из секреторных гранул, а количество инсулиновой мРНК в клетке возрастает в результате активации транскрипции и стабилизации мРНК. Активация транскрипции требует образования метаболитов глюкозы на стадиях гликолиза. Синтез и секреция инсулина не являются прочно сопряженными процессами. Например, при отсутствии ионов Са2+ в среде глюкоза не стимулирует секрецию инсулина, в то время как синтез активируется. Глюкоза стимулирует синтез инсулиновой мРНК при продолжительной инкубации (2—72 ч). При инкубации в течение 1 ч сколько-нибудь существенного увеличения мРНК не происходит, в то же время включение меченых аминокислот в проинсулин возрастает в 10—20 раз. Актиномицин D (ингибитор транскрипции) при этом не подавляет синтез проинсулина. Из этого следует, что первоначальная стимуляция синтеза (в течение примерно 20 мин после добавления глюкозы) происходит с использованием предсуществующей мРНК и регулируется на уровне трансляции.
Секреция инсулина и С-пептида происходит путем экзоцитоза. Инсулин в растворе легко образует олигомерные агрегаты, преимущественно димеры и гексамеры; ионы Zn2+ способствуют такой агрегации. В такой форме инсулин находится в секреторных гранулах. После секреции содержимого гранул в кровь олигомеры распадаются.
Глюкоза, аминокислоты (особенно аргинин и лизин), кетоновые тела и жирные кислоты в физиологических концентрациях стимулируют секрецию инсулина, причем стимуляция аминокислотами, кетоновыми телами и жирными кислотами проявляется при определенной (субстимулирующей) концентрации глюкозы. Лактат, пируват, глицерин такого влияния не оказывают. Глюкоза является главным регулятором секреции инсулина.
На рис. 5 показано изменение концентрации инсулина в крови человека после приема пищи. Одновременно со стимуляцией β-клеток к секреции инсулина происходит ингибирование секреции глюкагона из α-клеток панкреатических островков.
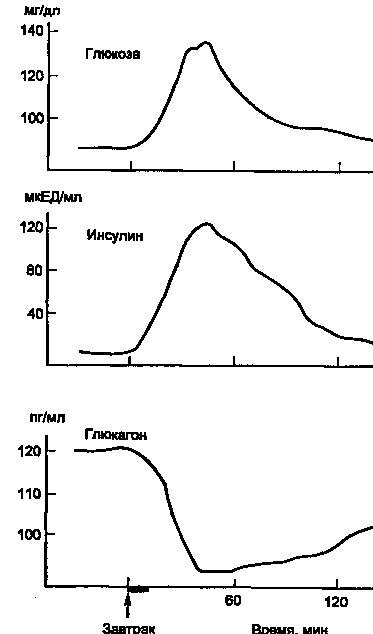
Рисунок 5. Изменение концентрации в крови глюкозы, инсулина и глюкагона после приема пищи (1 ЕД инсулина содержит 0,4081 мг белка инсулина).
Время полураспада инсулина в крови составляет 3—10 мин, а С-пептида около 30 мин. Кровь при однократном прохождении через печень теряет до 60 % инсулина. В почках задерживается до 40 % инсулина, содержащегося в протекающей через почки крови, причем в клубочках инсулин фильтруется, а затем наряду с другими белками первичной мочи (альбумин, гемоглобин и др.) реабсорбируется и разрушается в клетках проксимальных канальцев нефрона.
Регуляция секреции инсулина зависит от глюкозосенсорной системы β-клеток, обеспечивающей пропорциональность между концентрацией глюкозы в крови и секрецией инсулина. Потребление глюкозы β-клетками происходит при участии ГЛЮТ-1 (основной переносчик глюкозы в β-клетках человека) и, возможно, ГЛЮТ-2. Эта ступень не является лимитирующей: концентрация глюкозы в клетке быстро уравнивается с концентрацией в крови. В β-клетках глюкоза превращается в глюкозо-6-фосфат глюкокиназой (гексокиназой IV, как и в глюкозосинтезирующих органах — печени, почках), имеющей высокую Км для глюкозы — 12 мМ (Км гексокиназ I, II и III от 0,2 до 1,2 мМ). Вследствие этого скорость фосфорилирования глюкозы практически линейно зависит от ее концентрации в крови. Кроме того, глюкокиназа в Р-клетках — лимитирующее звено гликолиза. Поэтому глюкокиназа — вероятно, основной (но не единственный) элемент глюкозосенсорной системы β-клеток. Мутации глюкокиназы приводят к развитию одной из форм сахарного диабета диабету I типа у взрослых (MODY).
Специфический ингибитор глюкокиназы манногептулоза подавляет стимуляцию глюкозой синтеза и секреции инсулина. Это указывает на то, что молекулы, непосредственно регулирующие синтез и секрецию инсулина, образуются в результате метаболизма глюкозы. Природа этих молекул неизвестна. Согласно имеющимся представлениям, роль такой молекулы может выполнять АТР (точнее, отношение ([ATP]/[ADP]). Гипотеза обосновывается тем, что секреция инсулина стимулируется только метаболизируемыми веществами — источниками энергии. Например, глюкоза и глицеральдегид стимулируют секрецию пропорционально скорости их метаболизма. Глицерин не метаболизируется в Р-клетках (низкая активность глицеролкиназы) и не стимулирует секрецию инсулина. Однако после обработки рекомбинантным аденовирусом, содержащим бактериальный ген глицеролкиназы, клетки приобретают способность отвечать на глицерин секрецией инсулина в такой же мере, как и на глюкозу.
Есть указание на участие в регуляции секреции инсулина не только гликолиза, но и митохондриальных процессов. В частности, существенное значение могут иметь анаплеротические (восполняющие, компенсирующие) реакции: пируват → оксалоацетат, глутамат → α-кетоглутарат.. Эти реакции увеличивают количество компонентов цитратного цикла, а следовательно, и его мощность. Стимулированная глюкозой секреция инсулина усиливается некоторыми аминокислотами, жирными кислотами, кетоновыми телами; таким образом, в стимуляции секреции участвует не только глюкоза, но все основные энергоносители. Следовательно, количество секретируемого инсулина пропорционально энергетической ценности потребляемой пищи. Окисление основных энергоносителей в цикле лимонной кислоты, усиленном анаплеротическими реакциями, может быстро привести к изменению отношений ATP/ADP и NADH/NAD+ в клетке. Изменение концентрации этих веществ в свою очередь приводит к появлению вторых вестников сигнала (возможно, ионов Са2+, сАМР, диацилглицерола, инозитол-3-фосфата), которые включают процесс экзоцитоза инсулиновых гранул.
Механизмы активации экзоцитоза остаются неясными. Ряд экспериментальных данных указывает на участие Са2+/кальмо-дулинзависимой протеинкиназы (СаМПК), а также полифункциональной СаМПК II, которая найдена в панкреатических островках крысы и активируется глюкозой.
Глюкокиназа — основной элемент глюкозосенсорного механизма Р-клеток; она имеется также и в β-клетках, а гликолиз ускоряется пропорционально внеклеточной концентрации глюкозы и в тех, и в других клетках. Между тем секреция гормона (инсулина и глюкагона соответственно) стимулируется глюкозой в β-клетках и подавляется в β-клетках. Возможно, это связано с тем, что в β-клетках в отличие от а-клеток очень высокая активность пируваткарбоксилазы (анаплеротический фермент), сравнимая с активностью в клетках, для которых характерен глюконеогенез (печень, почки). При этом наблюдается пропорциональность между увеличением концентрации цитрата и малата в клетках и секрецией инсулина. Можно думать, что какие-то метаболиты этих путей или связанная с ними активация пируватмалатного челночного механизма участвует в сопряжении стимула с секрецией инсулина.
Популяция β-клеток в панкреатических островках неоднородна. В частности, есть клетки с различной чувствительностью к глюкозе. Это еще один элемент глюкозосенсорного механизма: при высокой концентрации глюкозы увеличивается число клеток, секретирующих инсулин.
НАРУШЕНИя метаболизма углеводов и липидов при сахарном диабете
При сахарном диабете инсулин-глюкагоновый индекс снижен. Это связано не только с уменьшением секреции инсулина, но и с увеличением секреции глюкагона (инсулин ингибирует секрецию глюкагона). В результате оказывается ослабленной стимуляция процессов складирования и усиленной стимуляция мобилизации запасов, причем настолько, что печень, мышцы, жировая ткань даже после приема пищи функционируют в режиме постабсорбтивного состояния (см. рис. 2). При этом продукты переваривания, а также их метаболиты, вместо того чтобы складироваться в форме гликогена и жиров, циркулируют в крови. Вероятно, в какой-то мере происходят и затратные циклические процессы типа одновременно протекающих гликолиза и глюконеогенеза или синтеза и распада жиров и т.п.
Для всех форм сахарного диабета характерна сниженная толерантность к глюкозе, т.е. гиперглюкоземия после приема пищи или даже натощак.
Основные причины гиперглюкоземии:
- потребление глюкозы мышцами и жировой тканью ограничено, поскольку в отсутствие инсулина ГЛЮТ-4 не экспонирован на поверхности миоцитов и адипоцитов.
Следовательно, глюкоза не используется для запасания в форме гликогена в мышцах и в форме жиров — в жировой ткани;
- в печени глюкоза не используется для запасания в форме гликогена, поскольку при низкой концентрации инсулина и высокой глюкагона гликогенсинтаза находится в фосфорилированной неактивной форме;
- в печени глюкоза не используется и для синтеза жиров: ферменты гликолиза и пируватдегидрогеназа находятся в неактивной форме и, следовательно, заторможено превращение глюкозы в ацетил-СоА, необходимый для синтеза жирных кислот;
- путь глюконеогенеза при низкой концентрации инсулина и высокой глюкагона активирован и возможен синтез глюкозы из аминокислот и глицерина.
Другим характерным признаком сахарного диабета является повышенная концентрация в крови липопротеинов (главным образом ЛОНП), свободных жирных кислот и, главное, кетоновых тел. Это связано с тем, что пищевые жиры не депонируются в жировой ткани, поскольку сАМР-зависимая липаза адипоцитов находится в фосфорилированной (активной) форме. Отсюда и повышенное содержание свободных жирных кислот в крови. Жирные кислоты поглощаются печенью, часть их превращается в адипоцитах в триацилглицерины, которые в составе ЛОНП секретируются в кровь. Другая часть жирных кислот вступает в путь β-окисления в митохондриях печени, и образующийся ацетил-СоА используется для синтеза кетоновых тел.
КОМАТОЗНЫЕ СОСТОЯНИЯ (ОСТРЫЕ ОСЛОЖНЕНИЯ) ПРИ ДИАБЕТЕ КАК РЕЗУЛЬТАТ НАРУШЕНИЯ ОБМЕНА ГЛЮКОЗЫ И ЖИРОВ
При сахарном диабете возможны три основные формы коматозных состояний: кетоацидотическая кома с абсолютной инсулиновой недостаточностью; гиперосмолярная кома с умеренной недостаточностью инсулина; лактатацидотическая кома с выраженной гипоксией, сепсисом, сердечно-сосудистым шоком. Кроме того, при инсулинотерапии может быть гипогликемическая кома, связанная с передозировкой инсулина. Первые три состояния могут развиться не только при сахарном диабете, но и при действии многих других факторов (токсических, инфекционных и др.).
Три основные формы коматозного состояния практически никогда не встречаются по отдельности. Обычно преобладают проявления какой-нибудь одной из форм (часто гиперосмолярной), что и дает повод для выделения основных форм.
Первичной причиной кетоацидоза является инсулиновая недостаточность: в период комы С-пептид и иммунореактивный инсулин (ИРИ) в крови не определяются. Гипергликемия отмечается всегда (20—30 ммоль/л, иногда более). Ацидоз при диабетической коме—это следствие накопления органических кислот: кетоновых тел, а также лактата и пирувата. Концентрация кетоновых тел достигает 2 ммоль/мл (в 200 раз больше нормы); она повышается не только вследствие синтеза в печени, но и потому, что снижается экскреция кетоновых тел в связи с олигурией и анурией, которая часто бывает при коме. Снижение рН крови до 7 и ниже (норма 7,4) наблюдается всегда.
Развивается дегидратация: дефицит воды может быть до 10 % от общей массы тела. Количество циркулирующей жидкости уменьшается на 25—30 %, в результате чего снижается артериальное давление.
Кислородное и энергетическое голодание миокарда, уменьшение объема крови ведут к сердечно-сосудистой недостаточности. Возможны повышение свертываемости крови, инфаркт миокарда, инфаркты паренхиматозных органов, инсульт, периферические тромбозы.
Диабетическая кома развивается медленно, в течение нескольких дней, иногда может возникнуть за несколько часов. Появляются тошнота, рвота, черты лица заостряются, глаза западают, нарастают безучастность к окружающему, заторможенность, переходящая в глубокую кому (полностью выключенное сознание, отсутствие рефлексов, атония мышц и др.). В помещении, где находится больной, ощущается запах ацетона. Артериальное давление снижено, почти всегда наблюдается олигурия или анурия. Диабетическая кома требует немедленного проведения следующих мероприятий: 1) ликвидация инсулиновой недостаточности путем введения инсулина в дозах, обеспечивающих постепенное снижение концентрации глюкозы в крови до уровня, близкого к нормальному; 2) регидратация организма путем введения жидкости; 3) восстановление нормального солевого состава и рН жидкостей организма путем введения соответствующих солевых растворов; 4) восстановление запасов гликогена в организме.
Проявления комы обычно ликвидируются в течение 2—3 дней при непрерывно продолжающемся лечении, причем лечение в начальные часы имеет решающее значение для больного.
До развития методов лечения диабета инсулином больные умирали вскоре после начала болезни от диабетической комы. Однако и в настоящее время кома наблюдается нередко. В частности, первое проявление болезни в 15—30 % случаев сопровождается кетоацидозом и комой. Смертность от диабетической комы остается высокой — от 1 до 30 %. Основной причиной смерти больных диабетом в настоящее время являются поздние осложнения.
ГЛИКозилИРОВАНИЕ БЕЛКОВ ОДНА ИЗ ГЛАВНЫХ ПРИЧИН ПОЗДНИХ ОСЛОЖНЕНИЙ САХАРНОГО ДИАБЕТА
Поздние осложнения сахарного диабета связаны прежде всего с повреждением кровеносных сосудов (диабетические ангиопатии). Основной механизм повреждения тканей — гликирование (гликозилирование) белков — не ферментативная реакция глюкозы со свободными аминогруппами белковой молекулы (Лиз, Арг, N-концевая аминокислота):
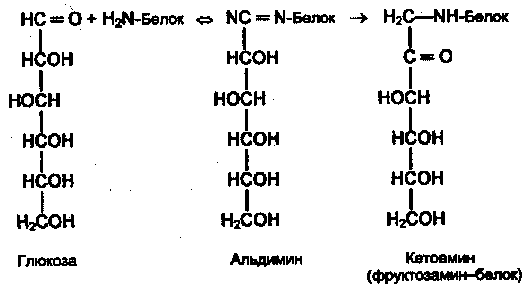
Вначале образуется нестабильная альдиминовая группировка, которая может превращаться в ряд других, более стабильных соединений («ранние продукты гликозилирования»). Понятно, что функции белка могут быть нарушены в результате изменения заряда белковой молекулы, ее конформации или блокирования активного центра. Гликозилирование — медленная реакция, в тканях здоровых людей обнаруживаются лишь небольшие количества гликозилированных белков. При гипергликемии реакция существенно ускоряется. Например, у больных диабетом в состоянии гипергликемии содержание одного из гликозилированных гемоглобинов HbAlc — в течение 2—3 нед увеличивается в 2—3 раза. Степень гликозилирования разных белков неодинакова; в основном она зависит от скорости обновления данного белка. В медленно обменивающихся белках накапливается больше модифицированных аминогрупп. Кроме того, в таких белках происходят дальнейшие изменения углеводных остатков: перестройка структуры, окислительные превращения, в результате которых образуются разнообразные «поздние продукты гликозилирования» (ППГ), часто коричневого цвета, флюоресцирующие, и некоторые из них обладают высокой реакционной активностью и способностью дополнительно повреждать белки, в том числе образовывать поперечные сшивки между молекулами белков. К медленно обменивающимся белкам относятся многие белки соединительно-тканных образований, межклеточного матрикса, базальных мембран. К тому же белки этих структур непосредственно контактируют с межклеточной жидкостью, в которой концентрация глюкозы такая же, как в крови (в клетках она обычно гораздо ниже в результате использования глюкозы в метаболических процессах). В этих структурах ППГ накапливаются с возрастом, накопление сильно ускоряется при сахарном диабете.
ППГ-белки могут гидролизоваться макрофагами (с участием ППГ-рецепторов) или межклеточными протеолитическими системами с образованием ППГ-пептидов, часто длиной около 30 аминокислотных остатков. ППГ-белки, особенно образующиеся в результате их гидролиза ППГ-пептиды, попадают и в кровоток. Концентрация ППГ-пептидов в крови резко повышается при почечной недостаточности разного происхождения, в том числе при диабетической нефропатии. Это связано с тем, что элиминация ППГ-пептидов происходит с участием почек: ППГ-пептиды фильтруются в клубочках, реабсорбируются клетками проксимальных канальцев и катаболизи-руются в лизосомах этих клеток.
В экспериментах на крысах показано, что введение ППГ-белков в кровь приводит к ковалентному связыванию этих белков с белками межклеточного матрикса во многих тканях и к появлению структурных и функциональных нарушений, сходных с теми, которые бывают при сахарном диабете.
ППГ проявляют многообразную биологическую активность: повышают проницаемость эндотелиальных клеток, соединяются с рецепторами макрофагов, эндотелиальных и мезангиальных клеток, активируют макрофаги к секреции цитокинов (рецепторным путем), подавляют образование NО и соответственно ингибируют расширение сосудов, усиливают окисление ЛНП. В крови больных диабетом обнаруживаются антитела к ППГ-пептидам.
Нарушение белкового обмена
Выраженный дефицит инсулина сопровождается отрицательным азотистым балансом и резким белковым истощением. При ювенильном инсулин-зависимом диабете частым осложнением в случае некомпенсированного заболевания является задержка роста. Такие нарушения не вызывают удивления, так как инсулин, если он присутствует в нормальных количествах стимулирует синтез белка и поглощение аминокислот мышцами и тормозит расход белка и высвобождение аминокислот мышцами. При инсулин-зависимом диабете изменяется содержание аминокислот в крови, их поглощение печенью и высвобождение мышцами. Отмечают снижение концентрации аланина в плазме крови и повышение концентрации других аминокислот. Несмотря на снижение уровня аланина в плазме, поглощение этой глюкогенной аминокислоты и других предшественников глюкозы печенью значительно увеличивается (в 2 – 10 раз).
У больных диабетом количество азотистых продуктов в мышце после приема белковой пищи восстанавливается труднее, чем в норме. Кроме того, аминокислоты, захваченные мышечной тканью, не включаются в белок, а преимущественно распадаются. При распаде аминокислот образуется аммиак, мочевина и другие продукты. В связи с этим при не леченном или декомпенсированном сахарном диабете возникают гиперазотемии с последующей гиперазотурией. Последняя обусловлена усиленным образованием аммиака как в печени, так и в почках из глутамина.
Список литературы:
1. Дедов И.И. Сахарный диабет в Российской Федерации: проблемы и пути решения//Сахарный диабет. - 1998. - № 1. - с. 7-18.
2.
Cheatham В.,
Kahn C.R. Insulin action and insulin signaling
network//Endocrine Rev. — 1995. — Vol. 16. — P. 17—142.
3. Ленинджер А. Основы биохимии. М.: Мир, 1985. – Т 2. 736 с.
4. Марри Р., Греннер Д., Мейерс П., Родуэл В. Биохимия человека. – M., Мир, 1993. – Т 2, 416 с.
5. Албертс Б., Брей Д., Льюис Д., Рэфф М., Робертс К., Уотсон. Д. Молекулярная биология клетки. – М., Мир, 1987. Т 3, 296 с.
6. Биохимические основы патологических процессов. // Под. ред. Северина Е.С. – М.: Медицина, 2000. 304 с.
7. Германюк Е.Л. Гликозилированные белки крови при сахарном диабете. // Клиническая медицина, 1982, Т 60, №10, с. 17 – 21.
© 2009 База Рефератов