
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Клинические признаки воспаления
Реферат: Клинические признаки воспаления
РЕАКЦИЯ ВОСПАЛЕНИЯ - универсальная защитная реакция организма на действие различных патогенных факторов (механических, физических, химических, биологических и др.), благодаря которой происходит обезвреживание и уничтожение вызвавших повреждение факторов. Проникшие патогены индуцируют комплексную реакцию воспаления, которая обеспечивает привлечение лейкоцитов и растворимых компонентов плазмы в очаги инфекции, что приводит к локализации и уничтожению микрорганизмов в области их внедрения.
КЛИНИЧЕСКИЕ ПРИЗНАКИ ВОСПАЛЕНИЯ
· rubor (краснота),
· tumor (опухоль, припухлость),
· calor (жар),
· dolor (боль),
· functio laesa (нарушение функции).
СТАДИИ ВОСПАЛЕНИЯ
В классической патологии было принято выделять три стадии воспаления:
- альтерацию,
- экссудацию,
- пролиферацию.
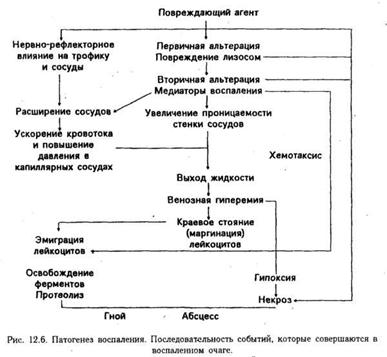
Такое деление сохраняется и в настоящее время. Однако новые исследования и новые факты показали, что указанные стадии не монолитны, между ними нет четких границ (например, альтерация может быть максимально выражена на стадии гнойной экссудации, а нарушения микроциркуляции могут быть неодинаковыми в одно и то же время в различных участках очага воспаления). Поэтому в зависимости от процесса, преобладающего на определенном этапе воспаления, выделяют следующие стадии (рис.):
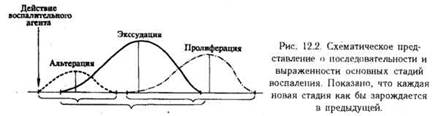
Стадия альтерации (повреждения):
· Первичная альтерация
· Вторичная альтерация
Стадия экссудации и эмиграции
Стадия пролиферации и репарации:
· Пролиферация
· Завершение воспаления
Первичная альтерация. Воспаление всегда начинается с повреждения ткани. После воздействия этиологического фактора в клетках происходят структурные, а также метаболические изменения.
Они бывают разными в зависимости от силы повреждения, от вида клеток (степени зрелости) и т.д.
Одни клетки гибнут, другие продолжают жить, третьи даже активируются.
Последние будут играть особую роль в дальнейшем.
Вторичная альтерация. Если первичная альтерация является результатом непосредственного действия воспалительного агента, то вторичная не зависит от него и может продолжаться и тогда, когда этот агент уже не оказывает влияния (например, при лучевом воздействии).
Этиологический фактор явился инициатором, пусковым механизмом процесса, а далее воспаление будет протекать по законам, свойственным ткани, органу, организму в целом.
Действие флогогенного агента проявляется, прежде всего, на клеточных мембранах, в том числе на лизосомах.
Это имеет далеко идущие последствия. Заключенные в лизосомах ферменты реактивны. Но как только лизосомы повреждаются и ферменты выходят наружу, они активируются и усугубляют, то разрушительное действие, которое оказал этиологический фактор. Можно сказать, что первичная альтерация — это повреждение, нанесенное со стороны, а вторичная альтерация — это самоповреждение.
Здесь следует отметить, что вторичная альтерация представляет собой не только повреждение и разрушение.
Некоторые клетки действительно гибнут, другие же не только продолжают жить, но и начинают вырабатывать биологически активные вещества, вовлекая в динамику воспаления иные клетки как в зоне воспаления, так и вне ее.
Клетки воспаления. Макрофаги. Установлено, что активированные макрофаги синтезируют особое вещество, получившее название интерлейкин-1 (ИЛ-1).
Оно выделяется макрофагами в среду и распространяется по всему организму, где оно находит свои мишени, которыми являются миоциты, синовиоциты, гепатоциты, костные клетки, лимфоциты, нейроциты.
По-видимому, на мембранах этих клеток имеются специфические рецепторы, благодаря которым ИЛ-1 действует именно на них, а не на другие клетки. Действие это стимулирующее и более всего изучено применительно к гепатоцитам и лимфоцитам.
Действие ИЛ-1 универсально, т.е. срабатывает при любом инфекционном (воспалительном) заболевании, причем в самом начале, и таким образом дает сигнал указанным органам включиться в воспалительный (инфекционный) процесс.
Есть основание предполагать, что характерные для ранних этапов заболевания симптомы (головная боль, боль в мышцах и суставах, сонливость, лихорадка, лейкоцитоз и увеличение содержания белков, в том числе иммуноглобулинов) объясняется именно действием ИЛ-1 (рис.).
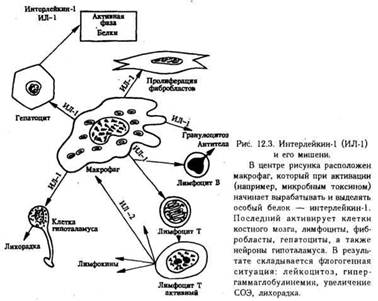
Роль макрофагов не ограничивается секрецией ИЛ-1. В этих клетках синтезируется еще целый ряд биологически активных веществ, каждое из которых делает свой вклад в воспаление. К ним относятся: эстеразы, протеазы и антипротеазы; лизосомальные гидролазы — коллагеназа, аластаза, лизоцим, α-макроглобулин; монокины ИЛ-1, колониестимулирующий фактор, фактор, стимулирующий рост фибробластов; антиинфекционные агенты — интерферон, трансферрин, транскобаламин; компоненты комплемента: С1, С2, СЗ, С4, С5, С6; дериваты арахидоновой кислоты: простагландин Е2, тромбоксан А2, лейкотриены. Не следует забывать также важнейшую функцию макрофагов — фагоцитоз.
Тучные клетки. Роль этих клеток в воспалении заключается в том, что они при повреждении выбрасывают содержащиеся в их гранулах гистамин и гепарин. А так как эти клетки в большом количестве располагаются по краям сосудов, то и действие указанных веществ проявится, прежде всего, на сосудах (гиперемия).
Макрофаги и лаброциты находятся в тканях постоянно (клетки-резиденты). Другие клетки воспаления проникают в зону воспаления со стороны (клетки-эмигранты). К ним относятся полиморфно-ядерные нейтрофилы, эозинофилы и лимфоциты.
Нейтрофилы. Главная функция этих клеток фагоцитоз. Они выселяются из костного мозга в кровь, эмигрируют из сосудов и в больших количествах скапливаются в воспаленной ткани.
И активное их размножение, и миграция, и фагоцитоз подвержены регулирующему влиянию биологически активных веществ (тканевых, системных, организменных). Действие их проявляется, однако, только тогда, когда на клетках имеются рецепторы, специфически реагирующие с медиатором воспаления: гистамином, адреналином, глюкокортикоидами, гамма-глобулинами и т.д.
В цитоплазме нейтрофилов имеется два типа гранул: первичные азурофильные (более крупные) — обычные лизосомы, вторичные, или специфические гранулы мельче, а главное, они содержат другой набор ферментных и неферментных веществ. В первичных гранулах содержатся кислые гидролазы, а кроме того, лизоцим, миелопероксидаза и катионные белки. Вторичные гранулы специфические; содержат щелочную фосфатазу, лактоферрин и лизоцим. Все это важно для понимания участия нейтрофилов в воспалении (см. ниже).
Эозинофилы. Роль эозинофилов в воспалении определяется рецепторами, расположенными на поверхности, и ферментами, находящимися внутри. На наружных мембранах имеются рецепторы для комплемента, иммунных комплексов, содержащих IgE, IgG. Из ферментов следует упомянуть гистаминазу и арилсульфатазу В. Важную роль играет большой катионный белок, который способен нейтрализовать гепарин, повреждать личинки ряда паразитов. Миграция и активация эозинофилов происходит под влиянием комплемента (С5а и С5—С7), пептидов тучных клеток, ПГД, веществ, продуцируемых гельминтами.
Тромбоциты. Роль тромбоцитов (кровяных пластинок) в воспалении состоит главным образом в том, что они имеют ближайшее отношение к микроциркуляции. Наверное, это самые постоянные и самые универсальные участники воспаления. В них содержатся вещества, влияющие на проницаемость сосудов, на их сократимость, на рост и размножение клеток, а главное — на свертываемость крови.
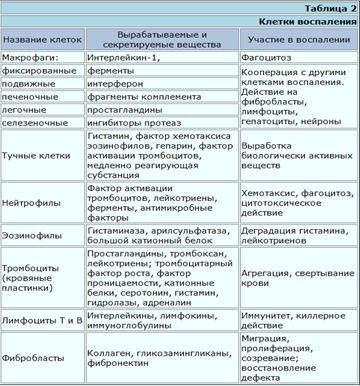
Лимфоциты. Эти клетки играют роль при любом воспалении, но особенно при иммунном.
Фибробласты. Действие фибробластов проявляется в последней стадии процесса, когда в очаге воспаления увеличивается число этих клеток, оживляется синтез в них коллагена и гликозамингликанов.
В суммарном виде данные о клетках воспаления представлены в табл. 2.
Медиаторы воспаления. Медиаторами воспаления называются биологически активные вещества, которые синтезируются в клетках или в жидкостях организма и оказывают непосредственное влияние на воспалительный процесс. Клеточные медиаторы были рассмотрены выше (см. табл. 2). Гуморальные медиаторы воспаления синтезируются в плазме и в тканевой жидкости в результате действия соответствующих ферментов. Первоначальной причиной появления (или увеличения количества) этих веществ является альтерация. Именно в результате повреждения клеток освобождаются и активируются лизосомальные ферменты, которые активируют другие ферменты, в том числе содержащиеся в плазме, в результате чего возникает целый ряд биохимических реакций. Поначалу они носят хаотический характер ("пожар обмена"), а продукты расщепления не имеют физиологического значения, нередко токсичны. Постепенно, однако, в этом процессе появляется определенный биологический смысл. Протеолитические ферменты расщепляют белки не до конца, а только до определенного этапа (ограниченный протеолиз), в результате чего образуются специфические вещества, действующие целенаправленно и ызывающие специфический патофизиологический эффект. Оказалось, что одни из них действуют преимущественно на сосуды, повышая их проницаемость, другие — на эмиграцию лейкоцитов, третьи — на размножение клеток. Первым обнаружил определенный "порядок" и закономерность в процессе воспаления В. Менкин. В воспалительном экссудате он выявил и индивидуализировал химические вещества и сопоставил с ними определенные слагаемые воспаления: гиперемию, лейкоцитоз, хемотаксис.
Одним из клеточных медиаторов воспаления является гистамин. Он содержится в гранулах тканевых базофилов (тучные клетки или лаброциты) в комплексе с гепарином и химазой в неактивной форме. В. свободном состоянии он оказывает расширяющее действие на мелкие сосуды (капилляры, венулы), увеличивая проницаемость их стенки. В малых дозах гистамин расширяет артериолы, в больших — суживает венулы. Выброс гистамина осуществляется вместе с выбросом в окружающую среду всех или части гранул тканевых базофилов при их дегрануляции. Этому может способствовать воздействие тепла, ионизирующего или ультрафиолетового излучения, растворов солей, кислот, белков, синтетических полимеров и мономеров, поверхностно-активных веществ. Дегрануляция всегда наблюдается при иммунных реакциях, т.е. при взаимодействии антигена с антителом на поверхности тканевых базофилов.
Другим клеточным медиатором воспаления является серотонин. У человека он содержится в тромбоцитах, хромаффинных клетках слизистой оболочки кишок, а также в некоторых нервных структурах. При разрушении клеток серотонин поступает в среду, вызывая повышение проницаемости сосудов.
Тканевые базофилы вырабатывают также гепарин, роль которого при воспалении заключается в том, что он препятствует образованию фибрина на внутренней оболочке капилляров, способствуя также увеличению проницаемости их стенки.
Лимфокины — вещества белковой природы, образующиеся в лимфоцитах, также относятся к медиаторам воспаления. Описано более десяти различных лимфокинов. При воспалении наибольшее значение имеют три из них: фактор, угнетающий эмиграцию макрофагоцитов, фактор, активирующий макрофагоциты, фактор хемотаксиса.
В клетках крови (лейкоцитах, тромбоцитах и др.) образуется еще одна группа веществ, играющих важную роль в динамике воспаления.
Это простагландины. Источником их образования являются фосфолипиды клеточных мембран. Нарушение строго упорядоченной структуры фосфолипидов в мембране делает их доступными действию фосфолипазы А2, в результате чего отщепляется арахидоновая кислота. С нее начинается каскад химических реакции, идущих в двух направлениях. Если на арахидоновую кислоту действует фермент циклоксигеназа, то в итоге образуются простагландины (ПГЕ2, ПГФ2, ПГИ2) или простациклины (ПГИ2), если же свою активность проявляет прежде всего липоксигеназа, то получаются лейкотриены. Дальнейшее превращение простагландинов происходит под влиянием тромбоксансинтетазы, в результате чего образуется тромбоксан А. Последний вызывает сужение сосудов, агрегацию тромбоцитов, тромбоз, отек, боль.
Другой путь биосинтеза простагландинов заключается в том, что под влиянием простациклинсинтетазы образуется простациклин (ПГИ2). Этот процесс совершается в эндотелиоцитах, где и находится указанный фермент. Он оказывает действие, противоположное тромбоксану: расширяет сосуды и подавляет агрегацию тромбоцитов.
Таким образом, арахидоновая кислота дает начало двум веществам с противоположным действием, причем выбор одного из путей биосинтеза, по-видимому, связан с состоянием эндотелия.
В неповрежденных эндотелиальных клетках содержится достаточно простациклинсинтетазы и весь ПГГ2 превращается в простациклин. Если же эндотелий поврежден, то этого фермента будет недоставать и потому часть ПГГ превращается в тромбоксан 2. Арахидоновый каскад представляет интерес еще и потому, что в ходе его образуются свободные радикалы, которые могут повреждать клеточные мембраны, в том числе и лизосом.
Лейкотриены оказывают хемотаксическое и хемокинетическое (нецеленаправленное движение) действие, повышают проницаемость, вызывают сокращение гладких мышц, индуцируют образование тромбоксанов.
К медиаторам воспаления относятся также циклические нуклеотиды, которые правильнее было бы назвать не медиаторами, а модуляторами, так как они не создают полной картины воспаления, а могут лишь в той или иной степени преобразовывать ее.
Циклические нуклеотиды обусловливают эффект действия других медиаторов, выделение клетками лизосомальных ферментов и др. Отмечено противоположно направленное действие цАМФ и цГМФ.
Так, первый подавляет выделение гистамина и лизосомальных ферментов, а второй, наоборот, способствует ему.
Из гуморальных медиаторов воспаления наибольшее значение имеют кинины — группа вазоактивных полипептидов, образующихся в результате каскада биохимических реакций, начинающихся с активации фактора Хагемана. Соприкосновение с поврежденной поверхностью или изменение внутренней среды (температура, рН) приводит к тому, что этот фактор становится активным и действует на находящийся в плазме прекалликреин, превращая его в калликреин.
Последний в свою очередь влияет на α2-глобулины, отщепляя от них полипептидную цепочку, состоящую из 9 (брадикинин) или 10 аминокислотных остатков (каллидин).
Плазменные кинины оказывают непосредственное влияние на тонус и проницаемость сосудистой стенки, вызывая расширение прекапиллярных артериол и увеличивая проницаемость стенки капилляров. Кроме того, они обусловливают типичные для воспаления зуд и боль.
Медиаторы калликреин-кининовой системы при воспалении влияют на реологические свойства крови, т.е. на ее способность находиться в жидком и текучем состоянии.
Активный фактор Хагемана может инициировать процессы кининообразования, гемокоагуляции и фибринолиза. Выпадение нитей фибрина и образование тромбов в зоне воспаления определенным образом связаны с состоянием калликреин-кининовой системы.
К гуморальным медиаторам воспаления относятся компоненты комплемента.
Известно, что последний является важным защитным фактором организма, но вместе с этим он может способствовать повреждению собственных тканей, что бывает при воспалении, особенно иммунном. Объясняется это тем, что из 9 компонентов комплемента 3 имеют ближайшее отношение к рассматриваемому процессу.
Так, комплемент С5 обладает способностью фиксироваться на сенсибилизированных и несенсибилизированных антителами клетках и разрушать их мембраны.
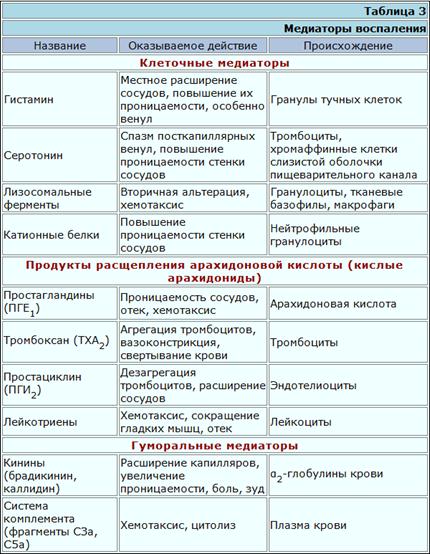
Фрагменты СЗа и С5а, а также трехмолекулярный комплекс С567 вызывают хемотаксис лейкоцитов. Наконец, клетки, нагруженные фрагментами С36, становятся объектом активного фагоцитоза. Характеристика основных медиаторов воспаления приведена в табл. 3.
Сосудистые расстройства. Экссудация и эмиграция. Воспаление характеризуется нарушением местного крово- и лимфообращения, прежде всего микроциркуляции. Микроциркуляцией принято называть движение крови в терминальном сосудистом русле (в артериолах, метартериолах, капиллярных сосудах и венулах), а также транспорт различных веществ через стенку этих сосудов. Микроциркуляцию удобно изучать с помощью опыта Конгейма (рис.).
При микроскопическом исследовании можно наблюдать, как сразу же после действия раздражителя (травма при извлечении кишки) возникает спазм артериол, который носит рефлекторный характер и скоро проходит. Вслед за этим развивается артериальная гиперемия. Она является результатом образования в воспаленном очаге большого количества вазоактивных веществ — медиаторов воспаления, которые расслабляют мышечные элементы стенки артериол и прекапилляров. Это приводит к увеличению притока артериальной крови, ускоряет ее движение, открывает ранее не функционировавшие капилляры, повышает в них давление. Кроме того, приводящие сосуды расширяются в результате паралича вазоконстрикторов, сдвига рН среды в сторону ацидоза, накопления ионов калия, снижения эластичности окружающей сосуды соединительной ткани.
Через 30 — 60 мин после начала эксперимента картина воспаления постепенно меняется: артериальная гиперемия сменяется венозной. При этом скорость движения крови уменьшается, меняется характер кровотока. Так, если при артериальной гиперемии клетки крови располагались главным образом в центре сосуда (осевой ток), а у стенок находились плазма и небольшое число лейкоцитов (плазматический ток), то теперь такое разделение нарушается. Изменяются реологические свойства крови. Она становится более густой и вязкой, эритроциты набухают, образуя агрегаты, т.е. беспорядочные скопления, которые медленно движутся или совсем останавливаются в сосудах малого диаметра.
Развитие венозной гиперемии объясняется действием ряда факторов, которые можно разделить на три группы:
· факторы крови,
· факторы сосудистой стенки,
· факторы окружающих тканей.
К факторам, связанным с кровью, относится краевое расположение лейкоцитов, набухание эритроцитов, выход жидкой части крови в воспаленную ткань и сгущение крови, образование тромбов вследствие активации фактора Хагемана, уменьшения содержания гепарина. Влияние факторов сосудистой стенки на венозную гиперемию проявляется набуханием эндотелия, в результате чего просвет мелких сосудов еще больше сужается. Измененные венулы теряют эластичность и становятся более податливыми сдавливающему действию инфильтрата. И, наконец, проявление тканевого фактора состоит в том, что отечная ткань, сдавливая вены и лимфатические сосуды, способствует развитию венозной гиперемии.
 С
развитием престатического состояния наблюдается маятникообразное движение
крови — во время систолы она движется от артерий к венам, во время диастолы
в противоположном направлении. Наконец, движение крови может полностью
прекратиться и развивается стаз, следствием которого могут быть необратимые
изменения клеток крови и тканей.
С
развитием престатического состояния наблюдается маятникообразное движение
крови — во время систолы она движется от артерий к венам, во время диастолы
в противоположном направлении. Наконец, движение крови может полностью
прекратиться и развивается стаз, следствием которого могут быть необратимые
изменения клеток крови и тканей.
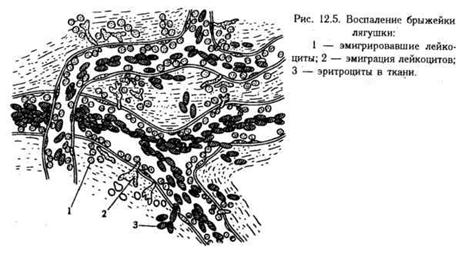
Одним из характерных признаков воспаления является экссудация и эмиграция лейкоцитов.
Экссудация — выход жидкой части крови, электролитов, белков и клеток из сосудов в ткани (рис.). Выход лейкоцитов (эмиграция) занимает в этом процессе особое место. Выходящая из сосудов жидкость (экссудат) пропитывает воспаленную ткань или сосредоточивается в полости, например в перикардиальной, в передней камере глаза и т.д.
Основной причиной экссудации является повышение проницаемости гистогематического барьера, т. е. сосудистой стенки, прежде всего капиллярных сосудов и венул. Исследования показали, что выход жидкости и растворенных в ней веществ осуществляется в местах соприкосновения эндотелиальных клеток. Щели между ними могут увеличиваться при расширении сосудов, а также, как полагают, при сокращении контрактильных структур и округлении эндотелиальных клеток. Кроме того, клетки эндотелия способны "заглатывать" мельчайшие капельки жидкости (микропиноцитоз), переправлять их на противоположный конец клетки и выбрасывать в близлежащую среду (экструзия).
Электронный микроскоп позволяет не только наблюдать эти микровезикулы, но измерить их и подсчитать число. Оказалось, что при воспалении происходит активизация микровезикулярного транспорта, что связано с затратой энергии. Об этом свидетельствует его остановка под влиянием ингибиторов образования макроэргических соединений.
Транспорт жидкости в ткани зависит от физико-химических изменений, происходящих по обе стороны сосудистой стенки. В связи с выходом белка его количество вне сосудов увеличивается, что способствует повышению онкотического давления. При этом происходит расщепление белковых и других крупных молекул на более мелкие. Гиперонкия и гиперосмия создают приток жидкости в воспаленную ткань. Этому способствует и повышение внутрисосудистого гидростатического давления в связи с изменениями кровообращения в очаге воспаления.
Экссудат отличается от транссудата тем, что содержит большее количество белков (более 2%). Если проницаемость стенки сосудов нарушена незначительно, то в экссудат, как правило, проникают альбумины и глобулины. При сильном нарушении проницаемости из плазмы в ткань поступает белок с большой молекулярной массой (фибриноген). При первичной, а затем и вторичной альтерации проницаемость сосудистой стенки увеличивается настолько, что через нее начинают проникать не только белки, но и клетки. При венозной гиперемии этому способствует . расположение лейкоцитов вдоль внутренней оболочки мелких сосудов и более или менее прочное их прикрепление к эндотелию (феномен краевого стояния лейкоцитов).
Прикрепление лейкоцитов к сосудистой стенке объясняется тем, что ее внутренняя оболочка при воспалении покрывается хлопьевидным слоем, в состав которого входит фибрин, гликозаминогликаны, гликопротеиды, сиаловые кислоты и др. На электрограммах этот слой имеет вид бахромы.
Эмиграция. Выход лейкоцитов из просвета сосудов через сосудистую стенку в окружающую ткань. Этот процесс совершается и в норме, но при воспалении приобретает гораздо большие масштабы. Смысл эмиграции состоит в том, чтобы в очаге воспаления скопилось достаточное число клеток, играющих роль в развитии воспаления (фагоцитоз и т.д.).
В настоящее время механизм эмиграции изучен довольно хорошо. С помощью микроскопа на живом объекте установлено, что лейкоцит пропускает между двумя эндотелиальными клетками свои псевдоподии, а затем и все тело. На электроннограммах видно, что лейкоциты выходят за пределы сосуда на стыке между эндотелиальными клетками. Это объясняется округлением эндотелиоцитов и увеличением интервалов между ними. После выхода лейкоцитов контакты восстанавливаются. Некоторые авторы допускают, что есть и второй путь эмиграции лейкоцитов — трансцеллюлярный, т. е. через цитоплазму эндотелиальных клеток. Однако в последнее время существование этого пути, по крайней мере, в норме, подвергается сомнению. После прохождения через слой эндотелия, лейкоциту предстоит преодолеть еще одно и, по-видимому, более значительное препятствие, а именно базальную мембрану. Она имеет толщину 40 60 нм и состоит из коллагеновых волокон и гомогенного вещества, богатого гликозаминогликанами. При прохождении через базальную мембрану полиморфно-ядерный лейкоцит атакует ее своими ферментами (эластаза, коллагеназа, гиалуронидаза). Они влияют на молекулярную структуру базальной мембраны, увеличивая ее проницаемость. Кроме ферментов, в этом случае определенную роль играют и содержащиеся в нейрофильных гранулоцитах катионные белки. Они действуют на коллоидное вещество мембраны, временно переводя его из геля в золь, увеличивая тем самым его проницаемость для клетки.
В эмиграции лейкоцитов в очаг воспаления наблюдается определенная очередность: сначала эмигрируют нейтрофильные гранулоциты, затем — моноциты и, наконец, — лимфоциты. Эту последовательность описал И. И. Мечников. Более позднее проникновение моноцитов объясняется их меньшей хемотаксической чувствительностью. После завершения воспалительного процесса в очаге наблюдается постепенное исчезновение клеток крови, начиная с тех лейкоцитов, которые появились раньше (нейтрофильные гранулоциты). Позже элиминируются лимфоциты и моноциты.
Клеточный состав экссудата в значительной степени зависит от этиологического фактора воспаления. Так, если воспаление вызвано гноеродными микробами (стафилококки, стрептококки), то в вышедшей жидкости преобладают нейтрофильные гранулоциты, если оно протекает на иммунной основе (аллергия) или вызвано паразитами (гельминты), то наблюдается множество эозинофильных гранулоцитов. При хроническом воспалении (туберкулез, сифилис) в экссудате содержится большое число мононуклеаров (лимфоциты, моноциты).
В очаге воспаления осуществляется активное движение лейкоцитов к химическим раздражителям, которыми могут быть продукты протеолиза тканей. Это явление описал И. И. Мечников и назвал его хемотаксис. Хемотаксис имеет значение на всех этапах эмиграции лейкоцитов, особенно во время движения в экстравазальном пространстве и в ткани, в которой отсутствуют сосуды (роговица). Если воспаление вызвано инфекционным агентом, то для хемотаксиса большое значение имеют продукты жизнедеятельности микроорганизмов, а также вещества, образующиеся в результате взаимодействия антигена и антитела.
В хемотаксисе лейкоцитов большое значение имеет система комплемента. Это, прежде всего компоненты комплемента СЗ и С5. Лейкотаксически активные компоненты комплемента СЗ и С5 могут образовываться под влиянием различных ферментов: трипсина, тромбина, плазмина.
Процесс эмиграции может не только стимулироваться, но и подавляться. Ингибиторы хемотаксиса вырабатываются активированными антигеном лимфоцитами. Понятно, что подвижность лейкоцитов будет уменьшаться, если на них подействовать такими ингибиторами обмена, как колхицин, пуромицин, актиномицин D, алкоголь.
В механизме движения лейкоцитов играют роль некоторые физико-химические факторы, например понижение поверхностного натяжения и выпячивание цитоплазмы в сторону раздражителя. Положительно заряженные макромолекулы могут уменьшать отрицательный заряд лейкоцитов и вызывать электростатическую неустойчивость их мембран. Это может привести к движению макромолекул (по типу укорочения — удлинение) как в цитолемме, так и в цитоплазме.
Фагоцитоз. В очаге воспаления главная функция лейкоцитов заключается в том, чтобы поглощать и переваривать инородные частицы (фагоцитоз). У одноклеточных организмов фагоцитоз служит для пищеварения, у высокоорганизованных эту функцию сохранили только некоторые клетки и она приобрела защитный характер. Все фагоцитирующие клетки И. И. Мечников разделил на микро- и макрофаги. Первые (полиморфно-ядерные лейкоциты) фагоцитируют микроорганизмы, вторые (моноциты, гистиоциты) поглощают и более крупные частицы, в том числе клетки и их обломки.
Различают четыре стадии фагоцитоза: приближение (хемотаксис), прилипание, поглощение, переваривание. Первая стадия (хемотаксис) была рассмотрена выше. Вторая стадия фагоцитоза — прилипание объясняется способностью фагоцитов образовывать тонкие цитоплазматические выпячивания, которые выбрасываются по направлению к объекту фагоцитоза и с помощью которых осуществляется прилипание. Определенное значение при этом имеет поверхностный заряд лейкоцитов. Лейкоциты с отрицательным зарядом лучше прилипают к объекту с положительным зарядом. Этому способствует модификация поверхности микроорганизмов, например, при фиксации на их поверхности антител (эффект опсонизации). Описано специальное вещество, которое стимулирует фагоцитоз. Это тафтсин (тетрапептид), который синтезируется в селезенке и активируется на клеточной мембране.
Третья стадия фагоцитоза поглощение объекта лейкоцитами может происходить двумя способами:
· контактирующий с объектом участок цитоплазмы втягивается внутрь клетки, а вместе с ним втягивается и объект;
· фагоцит прикасается к объекту своими длинными и тонкими псевдоподиями, а потом всем телом подтягивается в сторону объекта и обволакивает его. И в том, и в другом случае инородная частица окружена цитоплазматической мембраной и вовлечена внутрь клетки. В итоге образуется своеобразный мешочек с инородным телом (фагосома).
Четвертая стадия фагоцитоза — переваривание. Лизосома приближается к фагосоме, их мембраны сливаются, образуя единую вакуоль, в которой находятся поглощенная частица и лизосомальные ферменты (фаголизосома). В фаголизосомах устанавливается оптимальная для действия ферментов реакция (рН около 5,0) И начинается переваривание поглощенного объекта. На рис. 12.7 показан весь "арсенал", которым располагает активный микрофаг и который он пускает в ход на разных этапах фагоцитоза. Следует, однако, сказать, что одни ферменты не могут обеспечить достаточного киллерного действия. Эффективность фагоцитоза возрастает, когда в процесс подключается так называемая кислородная система. Как известно, в норме лейкоциты черпают энергию в основном вследствие гликолиза. При фагоцитозе повышается потребление кислорода, причем столь резкое, что его принято называть "респираторным взрывом" (рис.).
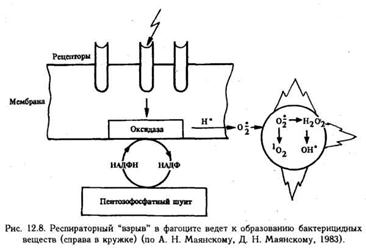
Смысл столь резкого (до 10 раз) повышения потребления кислорода состоит в том, что он используется для борьбы с микроорганизмами. Это происходит следующим образом. Заимствованный из среды, кислород активируется путем частичного восстановления. При этом образуется перекись водорода и так называемые свободные радикалы супероксидный анион — радикал О2•, синглетный кислород 1О2. Эти высокоактивные соединения вызывают перекисное окисление липидов, белков, нуклеиновых кислот, углеводов и при этом повреждают построенные из этих веществ клеточные структуры микроорганизмов. Особенно легко окисляются ненасыщенные жирные кислоты, входящие в состав клеточных мембран. Кислородный механизм пускается в ход, когда рецептор фагоцита приходит в контакт с объектом фагоцитоза. С этого момента начинается активация ферментов, которые переносят электроны на кислород, заимствуя их у восстанавливаемых молекул. Такими молекулами являются, прежде всего, НАДФН, которые под действием НАДФН-оксидаз окисляются до НАДФ, дефицит же НАДФН восполняется в результате окисления глюкозы в пентозофосфатном шунте. Эффективность действия такого окислителя, как перекись водорода, еще более возрастает, когда оно сочетается с действием миелопероксидазы и одного из галоидов (иона хлора или иода). Взаимодействие Н2О2, миелопероксидазы и хлора приводит к окислению ионов хлора и образованию гипохлорной кислоты, которая разрушает сульфгидрильные группы микробных ферментов, пептидные элементы клеточных мембран. Возможно, что эта реакция приобретает цепной характер, когда при окислении одних веществ образуются другие, сами являющиеся окислителями (например, альдегиды).
Конечно, в этой ситуации фагоцит и сам подвергается агрессивному действию названных веществ, но он обладает мощным механизмом, благодаря которому избыточного накопления активных форм кислорода не происходит. Защитную роль при этом играют, прежде всего, два фермента: глютатионпероксидаза и глютатионредуктаза, роль которых заключается в том, что первый переносит водород на окисленный глютатион, а второй — снимает этот водород и передает его на Н2О2, в результате чего образуются две молекулы воды.
Определенную роль играет каталаза, выводящая из клеток избыток перекиси водорода. Супероксидный анион обезвреживается особым ферментом — супероксиддисмутазой. У фагоцитов имеются и другие, не связанные с кислородом, механизмы борьбы с микроорганизмами. К ним относятся: лизоцим, разрушающий мембраны бактерий; лактоферрин, конкурирующий за ионы железа и, наконец, катионные белки, нарушающие структуру мембран микроорганизмов. Совместное действие механизмов обеих групп приводит к разрушению объекта фагоцитоза. Следует отметить, что фагоциты могут выделять в окружающую среду характерные для них ферментные и неферментные биологически активные вещества и тогда их действие происходит экстрацеллюлярно.
Нарушение обмена веществ в очаге воспаления. Интенсивность обмена веществ при воспалении, особенно в центре очага, повышается. Освобождающиеся из поврежденных лизосом ферменты гидролизуют находящиеся в очаге углеводы, белки, нуклеиновые кислоты, жиры. Продукты гидролиза подвергаются воздействию ферментов гликолиза, активность которых также повышается. Это относится и к ферментам аэробного окисления.
При изучении действия флогогенного агента (кротонового масла) на кожу в эксперименте было установлено, что потребление кислорода при этом повышается на 30 — 35 %. Однако это длится недолго — на протяжении 2 — 3 ч. Дальнейшая альтерация клеток сопровождается повреждением митохондрий — морфологического субстрата, на котором локализуются ферменты цикла Кребса и где осуществляется аэробное окисление и сопряженное с ним окислительное фосфорилирование. В связи с этим окисление еще более нарушается при почти неизменном гликолизе, что приводит к увеличению содержания молочной и трикарбоновых кислот (α-кетоглутаровой, яблочной, янтарной). Окисление при этом не завершается в цикле Кребса, снижается образование углекислоты, уменьшается дыхательный коэффициент.
Для характеристики метаболизма при воспалении издавна применяется термин "пожар обмена". Аналогия состоит не только в том, что обмен веществ в очаге воспаления резко повышен, но и в том, что "горение" происходит не до конца, а с образованием недоокисленных продуктов (полипетиды, жирные кислоты, кетоновые тела).
Следовательно, воспаление всегда начинается с усиления обмена веществ. Этим в значительной степени объясняется один из кардинальных признаков процесса — повышение температуры. В дальнейшем интенсивность метаболизма снижается, а вместе с этим изменяется и его направленность. Если сначала, т. е. в остром периоде воспаления, преобладают процессы распада, то в дальнейшем — процессы синтеза. Разграничить их во времени практически невозможно. Когда преобладают катаболические процессы, наблюдаются деполимеризация белково-гликозаминогликановых комплексов, распад белков, жиров и углеводов, появление свободных аминокислот, полипептидов, аминосахаров, уроновых кислот: Некоторые из образующихся веществ представляют особый интерес (кинины, простагландины), так как, включаясь в динамику воспаления, они придают ему определенный оттенок.
Анаболические процессы появляются очень рано, но преобладают на более поздних стадиях воспаления, когда проявляются восстановительные (репаративные) тенденции. В результате активирования определенных ферментов усиливается синтез ДНК и РНК, повышается активность гистиоцитов и фибробластов. В связи с повышением в них активности ферментов окислительно-восстановительных процессов активируются процессы окисления и окислительного фосфорилирования, увеличивается выход макроэргов.
Физико-химические изменения в очаге воспаления. Вследствие нарушения тканевого окисления и накопления в тканях недоокисленных продуктов развивается ацидоз. Сначала он компенсируется буферными механизмами, а затем становится декомпенсированным, в результате чего рН экссудата снижается. Концентрация ионов водорода тем выше, чем интенсивнее выражено воспаление. При остром абсцессе рН гноя может снизиться до 5,3. Наряду с повышенной кислотностью в воспаленной ткани повышается осмотическое давление, что является результатом усиления катаболических процессов: крупные молекулы расщепляются на более мелкие, их концентрация повышается. Увеличивается также содержание электролитов (ионов Na, К, Са). Концентрация ионов калия, освобождающегося из гибнущих клеток, в гнойном экссудате может достигать 256 511 ммоль/л (100 — 200 мг %), тогда как в нормальных тканях она не превышает 51,5 ммоль/л (20 мг %). Определение осмотического давления по снижению показателя точки замерзания экссудата показало, что депрессия составляет 0,6 0,8, что в пересчете на единицы измерения давления свидетельствует о повышении с 759 — 800 кПа (7,5 — 7,9 амт) в норме до 810 — 1114 кПа (8—11 атм) при воспалении (Шаде).
Ацидоз обусловливает набухание элементов соединительной ткани.
Повышение осмотического давления усиливает экссудацию и местный отек.
Этим объясняются главные признаки воспаления — появление припухлости и боли, наличие которой тоже в значительной степени объясняется натяжением ткани, возникающим при припухлости.
Пролиферация и завершение процесса. На этой стадии постепенно прекращаются разрушительные процессы и сменяются созидательными процессами. Прежде всего — это размножение клеток и возмещение возникшего ранее дефекта новообразованными клетками. Одновременно с размножением клеток и даже несколько опережая его, идет процесс активного погашения воспалительного процесса, что проявляется ингибицией ферментов, дезактивацией воспаления, расщеплением и выведением токсических продуктов. Активность клеток воспаления тормозится разными механизмами. Что касается ингибиторов, то в этом отношении важнейшую роль играет α2-макроглобулин (α2-М). Этот белок имеет широкий спектр действия. Он является главным ингибитором кининообразующих ферментов крови и таким образом устраняет их влияние: расширение и повышение проницаемости сосудов. Кроме того, он ингибирует большинство протеиназ из лейкоцитов, в том числе коллагеназу и эластазу и тем самым предохраняет от разрушения элементы соединительной ткани. Наконец, макроглобулин α2-М может связываться с мембранами нейтрофилов и таким образом тормозить их реакцию на СЗа и С5а (хемотаксис). В контроль за воспаление включаются и другие ингибиторы, в том числе альфа-антихимотрипсин (α1-АХ), который тормозит катепсин I и химотрипсин. Антитромбин III и α2-антиплазмин ингибируют сериновые ферменты и являются главными ингибиторами системы коагуляции, фибринолиза и комплемента.
В прекращении разрушительных явлений при воспалении важную роль играет устранение свободных радикалов. Внутри клеток свободные радикалы нейтрализует супероксиддисмутаза, а в экстрацеллюлярной фазе — церулоплазмин, катализируя реакцию НО2• + НО2• → Н2О2 + О2 .
Изменяется взаимоотношение между клетками. Они перестают вырабатывать одни медиаторы и начинают синтезировать другие. Теперь на тот же медиатор клетка может дать совсем другой ответ, потому что на ее поверхности появляются совсем другие рецепторы, а прежние проникают внутрь (интернализация).
Гистамин — типичный инициатор воспаления, но его эффект в конце воспаления может стать совсем иным, чем вначале. Оказалось, что это зависит от того, какие рецепторы "выставлены" на эффекторных клетках (например, на эндотелиоцитах) в данный момент. Если это Н1 то действие будет провоспалительное, а если Н2, то — противовоспалительное.
Полиморфно-ядерные нейтрофилы тоже подвергаются регулирующему влиянию тучных клеток. Гистамин, действуя на рецептор Н2, тормозит их хемотаксис, фагоцитарную активность, высвобождение лизосомальных протеаз. О сложных отношениях между клетками воспаления свидетельствует следующий факт. Известно, что тучные клетки при дегрануляции выделяют в среду два медиатора воспаления — гистамин и лейкотриен. В эозинофилах же содержатся два фермента, разрушающие эти медиаторы: гистаминаза и арилсульфатаза. Примечательно и то, что тучные клетки содержат еще одно вещество, привлекающее к ним эозинофилы. Это вещество называется фактором хемотаксиса эозинофилов. В итоге получается так, что эмигрирующие в зону воспаления эозинофилы разрушают медиаторы, выделенные тучными клетками, и тем самым гасят воспаление.
Для завершения процесса воспаления, особенно иммунного, большое значение имеет снижение активности лимфоцитов. В этом отношении гистамин тоже играет свою роль. Через рецепторы Н2 он тормозит секрецию лимфокинов, понижает митотическую активность лимфобластов, ограничивает активность Т-киллеров. Лимфокины инактивируются также и макрофагами через один из монокинов гистамина.
В инактивации клеток воспаления, кроме местных факторов, большую роль играют также и общие факторы, в том числе эндокринные. Гормон коры надпочечников кортизол тормозит синтез вазоактивных веществ в клетках, вызывает лимфопению, уменьшает число базофилов и эозинофилов. Кроме того, он стабилизирует мембраны лизосом.
На этом этапе воспаления меняются также свойства макрофагов: угнетается выработка интерлейкина-1. Что же касается фагоцитарной активности, то она к концу воспаления возрастает. Благодаря этому зона воспаления освобождается от некротизированных клеток, чуждых и токсических веществ.
После обезвреживания или локализации флогогенного фактора дальнейшие события направлены на ограничение зоны воспаления от остального организма, а затем замещения ее новой, здоровой тканью. Это осуществляется путем размножения оставшихся живых клеток (клетки резиденты), а также новых клеток из соседних зон (клетки эмигранты). Как миграция, так и пролиферация осуществляется за счет мезенхимальных клеток стромы. Размножаются стволовые клетки соединительной ткани — полибласты или лимфоидные клетки, адвентициальные и эндотелиальные клетки мелких сосудов, ретикулярные клетки лимфатических узлов. При дифференциации этих клеток образуются зрелые фибробласты, фиброциты, тучные и плазматические клетки, возникают новые капилляры. Все это совершается под контролем общих и местных факторов, которые могут как стимулировать, так и тормозить пролиферацию. К стимуляторам роста относятся специфические белки или пептиды, источником которых являются тромбоциты (тромбоцитарный фактор роста фибробластов). В лимфоцитах также образуется пептид, стимулирующий соединительную ткань, а в моноцитах стимулятор роста фибробластов (интерлейкин-1). Ряд веществ с таким же действием образуется не на тканевом, а на органном уровне: в гипофизе — фактор роста фибробластов, в печени — соматомедин, стимулирующий обмен веществ в фибробластах. Все эти вещества оказывают свое действие только в тех случаях, когда на клетках-мишенях (фибробластах, эндотелиоцитах) имеются соответствующие рецепторы.
Размножение клеток в нужных масштабах определяется не только стимуляторами, но и ингибиторами пролиферации. К последним относятся кейлоны. Механизм действия этих веществ хорошо изучен на эпителии. Эти вещества синтезируются и депонируются в зрелых клетках, которые слущиваются с кожи. Когда число этих клеток уменьшается, становится меньше и число кейлонов, а значит, снижается эффект торможения и деление клеток ускоряется. Воспалительный очаг характеризуется наличием малого числа зрелых клеток и, следовательно, низким содержанием кейлонов — депрессоров деления. Из гормональных ингибиторов роста следует назвать кортизон.
Действие стимуляторов размножения направлено на увеличение числа клеток и одновременно на усиление обмена веществ в них, их синтетических способностей. Это относится прежде всего к синтезу коллагена и гликозаминогликанов.
В конце воспаления, в деле его завершения решающую роль играют две клетки: фибробласт и эндотелиоцит. Два процесса совершаются в этот период: заселение зоны фибробластами и неоангиогенез, т. е. образование новых кровеносных и лимфатических сосудов. Для миграции фибробластов имеют значение следующие вещества: продукты распада коллагена, интерлейкин-1, фибронектин и один из лимфокинов. Размножаются фибробласты под влиянием пластиночного активирующего фактора (ПАФ-2) и соматомедина (синтезируется в гепатоцитах).
Зрелые фибробласты синтезируют два вещества, которые затем заполняют межклеточное пространство: коллаген и гликозаминогликаны. Клетки и эти два вещества могут составить основу того барьера, который изолирует очаг воспаления от остального организма.
Нити коллагена в межклеточном пространстве располагаются в определенном порядке и играют роль не только механическую (растяжение, сдавление), но также и структурную (порядок в расположении фибробластов, эпителиальных и других клеток) и, наконец, могут влиять на метаболизм клеток и даже на их геном.
Вместе с миграцией фибробластов в зону воспаления устремляются также эндотелиоциты. Их источником являются сохранившиеся капилляры. Эндотелиоциты активируются в отношении размножения, подвижности и обмена. Полагают, что направление движения (таксис) этих клеток объясняется градиентом кислорода (в сторону, где его меньше). Повышение метаболизма обеспечивает энергией (АТФ) сократительные элементы (актин) внутри клеток. О подвижности эндотелия свидетельствует тот факт, что при воспалении капилляры появляются в тканях, в которых в норме они не наблюдаются (роговица). Когда васкуляризация достигает максимального уровня, дальнейшее образование сосудов прекращается.
При небольших повреждениях тканей, при ранах, заживающих первичным натяжением, воспалительный процесс заканчивается полным восстановлением. При гибели больших массивов клеток дефект замещается соединительной тканью с последующим образованием рубца. Этим воспаление обычно и заканчивается. Однако в некоторых случаях наблюдается избыточное образование рубцовой ткани, которая может деформировать орган и нарушать его функцию. Это особенно опасно при воспалении клапанов сердца, мозговых оболочек и т. д.
СПИСОК ЛИТЕРАТУРЫ
Патологическая физиология [Учебник для студентов мед. вузов]
Н. Н. Зайко, Ю. В. Быць, А. В. Атаман и др. К.: "Логос", 1996
Интернет сайты:
http://ru.wikipedia.org/wiki/Воспаление
http://humbio.ru/Humbio/immunology/imm-gal/00061b26.htm
© 2009 База Рефератов