
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Координація періодичної діяльності гастродуоденальної зони при порушенні її функціонально-морфологічної цілісності
Реферат: Координація періодичної діяльності гастродуоденальної зони при порушенні її функціонально-морфологічної цілісності
ТАВРІЙСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ
імені В. І. ВЕРНАДСЬКОГО
ТРУШЕНКО Олександр Сергійович
УДК 612.3.002.2:616.329.002
КООРДИНАЦІЯ ПЕРІОДИЧНОЇ ДІЯЛЬНОСТІ ГАСТРОДУОДЕНАЛЬНОЇ ЗОНИ ПРИ ПОРУШЕННІ ЇЇ ФУНКЦІОНАЛЬНО-МОРФОЛОГІЧНОЇ ЦІЛІСНОСТІ
03.00.13 – фізіологія людини і тварин
АВТОРЕФЕРАТ
дисертації на здобуття наукового ступеня
кандидата біологічних наук
Сімферополь – 2008
Дисертацією рукопис
Робота виконана на кафедрі фізіології людини і тварин Дніпропетровського національного університету Міністерства освіти і науки України
Науковий керівник кандидат біологічних наук, професор
Мурзін Олексій Борисович
Дніпропетровський національний університет
професор кафедри фізіології людини і тварин
Офіційн опоненти: доктор біологічних наук, професор
Коренюк Іван Іванович
Таврійський національний університет
імен В. І. Вернадського,
професор кафедри фізіології людини і тварин та біофізики
кандидат біологічних наук, старший науковий співробітник
Харченко Микола Михайлович
НДІ фізіології імені академіка Петра Богача біологічного факультету Київського національного університету
імен Тараса Шевченка,
старший науковий співробітник відділу фармакофізіології
Захист відбудеться “____“_____________2008 р. о ______годині на засіданн спеціалізованої вченої ради К 52.051.04. у Таврійському національному університеті імені В. І. Вернадського за адресою:
95007, Крим, м. Сімферополь, проспект В.І. Вернадського, 4
З дисертацією можна ознайомитись у бібліотеці Таврійського національного університету ім. В. І. Вернадського за адресою:
95007, Україна, Крим, м. Сімферополь, проспект В. І. Вернадського, 4
Вчений секретар
спеціалізовано вченої ради,
кандидат біологічних наук, доцент Д. Р. Хусаінов
ЗАГАЛЬНА ХАРАКТЕРИСТИКА РОБОТИ
Актуальність теми. Ультрадіанні ритми шлунково-кишкового тракту одним із найважливіших механізмів, які регулюють та підтримують гомеостаз організму в кожний відповідний момент часу. Моторний компонент цієї складно періодичної активності, відомий як мігруючий моторний комплекс, сьогодні, як раніше, інтенсивно вивчається [Лебедев, 1987; Lee et al., 1995; Parkman et al., 1999; Tanaka et al., 2001; Кромин, 2002; Budzynski et al., 2002]. Взаємозв’язок моторних та секреторних патернів шлунково-кишкового тракту в умовах зміни циркадіанних ритмів не викликає сумнівів, проте на сьогодні актуальною залишається проблема перебудов ультрадіанних ритмів травної системи при різних змінах функціонального стану її регуляторних механізмів [Dalenbäck et al., 1998; Yang et al., 2004].
Незважаючи на велику кількість публікацій щодо ролі регуляторних механізмів травно системи у координації періодичної діяльності верхніх відділів шлунково-кишкового тракту, проблема адаптаційно-компенсаторних змін періодично моторно-секреторної активності в умовах різноспрямованих порушень місцевої нейроімуноендокринно регуляції зберігає актуальність зараз та загострюється у зв’язку з поширенням поєднаної патології гастродуоденальної зони [Xu et al., 2006] та остаточно не з’ясованою роллю нехолінергічної неадренергічної ланки в регуляції періодично активності гастродуоденальної зони за різних умов [Kato et al., 1998; Takeuchi et al., 1993; Beltran et al., 1999; Konturek et al., 1999; Takeuchi et al., 1995, 1997, 2000].
На сьогодні також залишається не вивченим стан шлункової секреції в динаміц моделювання дуоденогастрального рефлюксу in vivo, а також регулятивні зміни секреторної діяльності шлунка, особливо на перших етапах дії несприятливих факторів. Подібні порушення, з одного боку, супроводжуються стійкою десинхронізацією періодичної діяльності органів гастродуоденальної зони, а з ншої – розвитком специфічних адаптаційно-компенсаторних змін власне моторно-секреторного апарату гастродуоденальної зони [Tache et al., 2004; Лычкова, 2005], які також потребують подальшого поглибленого вивчення.
Окрім того, провідну роль у розвитку функціонально-морфологічних змін верхніх відділів шлунково-кишкового тракту за наявності дуоденогастрального рефлюксу відіграють зсуви регуляції моторної функції гастродуоденальної ділянки [McCallum, 1997]. Проте, з оглядом на відомий взаємозв’язок моторики та секреції цієї ділянки шлунково-кишкового тракту, практично невивченими залишаються особливості періодичної моторно-секреторної діяльност гастродуоденальної зони за умов розвитку дуоденогастрального рефлюксу in vivo.
Зв’язок роботи з науковими програмами, планами, темами. Дисертація пов’язана з фундаментальною науковими роботами: “Дослідження нервових механізмів керування, регуляції і зв’язку в організмі людини і тварин (№ державної реєстрації: 0195U023144), “Фундаментальні комплексні дослідження фізіологічних механізмів виникнення, розвитку та прояву серцево-судинних та шлунково-кишкових порушень” (№ державної реєстрації по Дніпропетровському національному університету: БЕФ-85), а також є частиною комплексно науково-дослідної роботи ДУ “Інститут гастроентерології” АМН України за темою Вивчити механізми функціональних та структурних змін верхнього відділу травного каналу при пептичній виразці дванадцятипалої кишки, поєднаній з рефлюксною хворобою та визначити шляхи їх корекції (клініко-експериментальне дослідження)”, шифр теми: ВН.25.01.002.04, № державної реєстрації: 0104U002440.
Об’єкт дослідження: Міоелектрична активність шлунка та дванадцятипалої кишки, а також показники секреторної діяльності шлунка при розвиткові вегетативного дисбалансу, дуоденогастрального рефлюксу у початковому стані та при блокад нітрергічних механізмів регуляції.
Предмет дослідження: Адаптивно-компенсаторні реакції періодичної моторно-секреторно активності гастродуоденальної зони при різних порушеннях функціонально-морфологічної цілісності верхніх відділів шлунково-кишкового тракту.
Методи дослідження: використані електрофізіологічні методи дослідження міоелектрично активності шлунка та дванадцятипалої кишки, методи збору та біохімічного аналізу шлункового соку, гістохімічні методи оцінки стану слизової оболонки шлунка та дванадцятипалої кишки, варіаційно-статистичні методи обробки експериментальних даних: розрахунок середньої та її похибки.
Мета дослідження: встановлення взаємозв’язків періодично моторної активності шлунка, дванадцятипалої кишки, діяльності секреторних залоз шлунка в умовах порушення морфофункціональної цілісності гастродуоденально зони.
Завдання наукового дослідження:
1) дослідити стан моторної активності шлунка при моделюванні вегетативного дисбалансу;
2) дослідити стан секреторного апарату шлунка при моделюванні вегетативного дисбалансу;
3) вивчити міоелектричну активність шлунка та дванадцятипалої кишки при моделюванні дуоденогастрального рефлюксу;
4) з’ясувати стан секреторної функції шлунка при моделюванні дуоденогастрального рефлюксу;
5) зіставити особливості моторної та секреторної функцій верхніх відділів шлунково-кишкового тракту при моделюванні дуоденогастрального рефлюксу;
6) дослідити стан моторної діяльності шлунка та дванадцятипалої кишки, а також секреторної активності шлунка при блокаді місцевих нітрергічних механізмів в нтактних тварин та при моделюванні дуоденогастрального рефлюксу;
7) з’ясувати участь нітрергічних механізмів у регуляції періодичної моторно-секреторно діяльності верхніх відділів шлунково-кишкового тракту в інтактному стані та при моделюванні дуоденогастрального рефлюксу.
Наукова новизна одержаних результатів. У роботі вперше проведено комплексне експериментальне дослідження моторно-секреторно діяльності гастродуоденальної зони на первинних етапах розвитку вегетативного дисбалансу та дуоденогастрального рефлюксу. Уперше встановлено, що при розвитку вегетативного дисбалансу зміни моторно-секреторних ритмів шлунка протікали в три стадії, що супроводжувалось зниженням функціональної здатності секреторних залоз шлунка, явищем «відмови в регуляції», якому передувала поява кисло шлункової секреції натще, а також розвитком дуоденогастрального рефлюксу.
Уперше показано, що на первинних етапах розвитку дуоденогастрального рефлюксу результатом адаптивно-компенсаторних перебудов періодичної діяльност гастродуоденальної зони було підвищення активності парієтальних та поверхнево-епітеліальних клітин слизової оболонки шлунка із супутнім пригніченням діяльності її головних клітин. Уперше виявлено, що при розвитку дуоденогастрального рефлюксу нітрергічна регуляція зберігає свій гальмівний вплив у відношенні скоротливої активності гастродуоденальної зони шляхом пригнічення міогенної пікової активності антрального відділу шлунка та цибулини дванадцятипалої кишки. При цьому відбувається перебудова місцево нейроендокринної регуляції гастродуоденальної зони, однією з механізмів яко протилежна зміна впливу NO-ергічної ланки у напряму діяльності парієтальних клітин шлунка. Новими є дані, які свідчать, що тривале введення блокаторів NO-синтази може призводити до порушень цілісності слизової оболонки шлунка та виникнення виразкових ушкоджень останньої.
Практичне значення одержаних результатів. Отримані в роботі дан можуть бути використані в науково-дослідній роботі при вивченн клініко-фізіологічних аспектів загального адаптаційного синдрому за умов розвитку стрес-реакції. Уперше запропоновано оригінальний нетравматичний спосіб збору шлункового соку у лабораторних щурів. Результати даної роботи можна враховувати в клінічній гастроентерології при вивченні змін періодично моторно-секреторної діяльності верхніх відділів шлунково-кишкового тракту при розвиткові дуоденогастроезофагеальної рефлюксної хвороби. Також основн положення роботи можуть бути врахованими при розробці нових функціональних проб та діагностичних критеріїв розвитку функціонально-морфологічних зсувів гастродуоденальної зони на перших етапах передонкологічних процесів у шлунці.
Практико-теоретична цінність дисертаційного дослідження підтверджується впровадженням результатів та пропозицій, які витікають з роботи, в навчально-практичний процес кафедри фізіології людини і тварин Дніпропетровського національного університету та в науково-дослідницьку діяльність ДУ “Інституту гастроентерології АМН України”.
Особистий внесок здобувача. Автором самостійно: проведено аналіз літературних джерел, виконані експерименти з вивчення міоелектрично секреторної активності шлунка та дванадцятипалої кишки у щурів; запропоновано та обґрунтовано оригінальний метод збору шлункового соку у щурів; проведено систематизацію та статистичний аналіз показників моторики та секрец гастродуоденальної зони завдяки відомим комп’ютерним програмам. У дисертації не використовувалися практичні розробки співавторів опублікованих робіт. У працях, написаних у співавторстві, здобувачеві належить: проведення (або участь у проведенні) експериментів, аналіз, систематизація та узагальнення результатів дослідження, та оформлення їх у вигляді статей.
Апробація результатів. Матеріали дисертації представлені у доповідях на конференціях ДНУ за підсумками науково-дослідних робіт у 2005–2007 рр.; на IV Міжнародній науково-практичній конференції “Динаміка наукових досліджень – 2005” (Дніпропетровськ, 2005 р.); на Міжнародній науковій конференції, приуроченій до 60-ліття новоствореної кафедри фізіології людини тварин Львівського університету ім. І. Франка (Львів, 2006 р.); на Всеукраїнській науково-практичній конференції «Вклад молодих вчених в розвиток медичної науки і практики» (Харків, 2006 р.); на VIII Міжнародній науково-практичній конференції «Людина і космос. Ракетно-космічна техніка» (Дніпропетровськ, 2006 р.); на ІІ Міжнародній конференції “Neural-humoral and cellular regulatory mechanisms of digestion processes” (Київ, 2005 р.); на XVII З’їзді українського фізіологічного товариства з міжнародною участю (Чернівці, 2006 р.); на IХ Міжнародній науково-практичній конференції «Людина і космос. 50-ліття космічної ери» (Дніпропетровськ, 2007 р.); на Всеукраїнській науковій конференції «Сучасні питання фізіології та медицини», присвяченій 85-річчю заснування кафедри фізіології людини і тварин та 100-річчю з дня народження П. Є. Моцного (Дніпропетровськ, 2007 р.).
Публікації. За темою дисертації опубліковано 10 робіт: 4 статті – у фахових журналах ВАК, 6 тезами доповідей з’їздів і конференцій.
Структура та обсяг дисертації. Дисертація складається зі вступу, огляду літератури, опису методів досліджень, двох розділів результатів досліджень, узагальнення результатів досліджень, висновків і списку використаних джерел. Дисертація викладена на 157 сторінках, містить 55 рисунків, 2 таблиці, список використаних джерел із 280 найменувань.
ОСНОВНИЙ ЗМІСТ
Методи дослідження. Всі експерименти були проведені відповідно до існуючих міжнародних правил використання лабораторних тварин у медико-біологічних дослідженнях. Для вирішення встановлених задач було проведено 6 серій експериментів (гострих, напівгострих та хронічних). У першій серії експериментів (N=15) вивчали міоелектричну активність шлунка собак у вихідному стані та в динаміц моделювання вегетативного дисбалансу. У другій серії (N=5) досліджували секреторну активність шлунка собак у вихідному стані та в динаміц моделювання вегетативного дисбалансу. У третій серії (N=22) досліджень вивчали міоелектричну активність шлунка та дванадцятипалої кишки нтактних щурів у вихідному стані та при блокуванні NO-синтази. У четвертій серії (N=24) досліджень вивчали секреторну функцію шлунка інтактних щурів у вихідному стані та при блокуванні NO-синтази. У п’ятій серії (N=27) вивчали міоелектричну активність шлунка та дванадцятипалої кишки при моделюванні дуоденогастрального рефлюксу відповідно на 3-й, 6-й та 12-й день у вихідному стані та при блокуванні NO-синтази. У шостій серії (N=36) досліджували шлункову секрецію у щурів при моделюванні дуоденогастрального рефлюксу відповідно на 3-й, 6-й та 12-й день у вихідному стані та за умов неселективного блокування NO-синтази. Моделювання вегетативного дисбалансу у собак проводили шляхом щоденних внутрішньом’язових ін’єкцій карбохоліну (0,01 мг/кг) за 15 хвилин до введення цинхофену (0,05 г/кг) натще per os протягом 30 діб. Для відтворення дуоденогастрального рефлюксу здійснювали щоденне введення жовчі per os натще із наступним впровадженням імобілізаційно-холодового стресу протягом однієї години (+4˚С) [Тарасенко и др., 2001, з модифік.]. Міоелектричну активність шлунка та дванадцятипалої кишки вивчали за допомогою платинових (у собак) та срібних (у щурів) біполярних електродів з використанням самописця WI-387G та системи «поліграф RM-86 Nihon Kohden–комп’ютер». Міоелектричну активність оцінювали за характером змін її складових (частоти та амплітуди повільних потенціалів, періоду основного (базального) електричного ритму, а також тривалості фаз мігруючого моторного комплексу). Неселективне блокування NO-синтази здійснювали за допомогою внутрішньочеревної ін’єкц NG-нітро-L-аргініну (L-NNA). Для вивчення секреторної функції шлунка собакам вживляли фістули за В. А. Басовим. Шлунковий сік у щурів збирали зондовим методом. Для з’ясування стану слизової оболонки гастродуоденально зони здійснювали макроскопічні та гістохімічні дослідження відповідних ділянок слизової оболонки шлунка та дванадцятипалої кишки із забарвленням гематоксилін-еозином, альциановим синім, та ШИК-реакцію за загальноприйнятими методиками [Крышень и др., 1978]. Активність NO-синтаз в секреторних залозах шлунка визначали за допомогою NADPH-діафоразної реакції [Shyamal et al., 2001]. Статистичну обробку отриманих результатів проводили за допомогою пакету програм Microsoft Excel та Origin 6.1 Pro з використанням t-критерію Стьюдента, а також непараметричних критеріїв Вілкоксона-Манна-Уітні для порівняння рядів даних з малою кількістю значень (N) [Гланц, 1999]. Досліджувані співвідношення вважали достовірними при Р<0,05.
Результати дослідження. Міоелектрична активність шлунка при моделюванні вегетативного дисбалансу. Моделювання вегетативного дисбалансу супроводжувалося розвитком виразкових ушкоджень слизової оболонки шлунка, як розташовувалися в місці переходу інтрамедіальної зони шлунка в препілоричну, або ж на ділянці воротаря на малій кривині шлунка. Якщо в інтактному стан міоелектрична активність шлунка складалася з 3-х фаз: фази I (60-90 хв), фази II (20–30 хв) і фази III (5-7 хв), то на 3–5-й день експерименту спостерігалося збільшення тривалості II-III фаз на 150,0–200,0% (до 40 хв), а тривалість I фази зменшилася на 30,0–40,0% (до 40–55 хв, P<0,01). Треба зазначити, що на 7–10-й день дослідження можна було спостерігати розповсюдження повільних хвиль міоелектричної активності в ретроградному напрямку, що передувало закиданню дуоденального вмісту в шлунок. У подальшому – на 15–35-й день – такий стан моторної активності супроводжувався антиперистальтичними хвилями, які часто закінчувалися блювотою. На ранніх етапах моделювання вегетативного дисбалансу відзначалася тенденція до зростання частоти повільних хвиль основного електричного ритму шлунка – до 10-го дня (рис. 1, А), при цьому період основного електричного ритму зменшувався (P<0,05).
|
А |
Б |
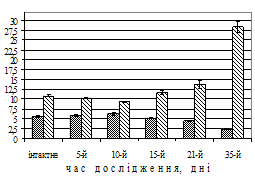
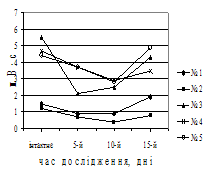
Рис. 1. Динаміка частоти (темні стовпчики) повільних потенціалів, періоду основного електричного ритму (світлі стовпчики) антрального відділу шлунка (А) та моторного індексу в ряді відведень шлунка собак (Б) при моделюванні вегетативного дисбалансу (зліва на рис. Б – схема розташування та номери відведень). По вісі абсцис – час від початку досліду, день; по вісі ординат – частота повільних потенціалів, за хв, період основного електричного ритму, с, значення моторного індексу, мВ∙с (M±m (А), M (Б), N=15 (інтактне), N=5 (дослід)).
У подальшому – на 15-й день – навпаки, зменшилася частота повільних потенціалів та зріс період основного електричного ритму майже в 3 рази (P<0,01). Треба зазначити, що вже з 3–5-го дня відзначалося зниження моторного індексу, особливо в перехідній зоні між антральним та фундальним відділами шлунка (рис. 1, Б). Неузгодженість у діяльності між фундальним та антральним відділами шлунка за даними моторного індексу можна було виявити саме до 15–20-го дня дослідження.
Секреторна діяльність шлунка при моделюванні вегетативного дисбалансу. Дослідження другої серії виявили, що вже на 10-й день у більшості тварин поряд із підвищенням слизевиділення натще спостерігалося виділення кислого соку (рис. 2, А). При цьому тривалість та інтенсивність останнього мали індивідуальні особливості у кожної тварини. У шлунковому вміст визначався пепсин, максимальна кількість якого припала на 21-й день. Починаючи з 10-го дня можна
|
А |
Б |
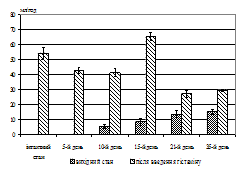
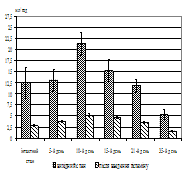
Рис. 2. Зміни дебіту базальної кислої шлункової секреції, об’єму кислої шлункової секреції (А) та дебіту слизу (Б) у собак після введення гістаміну в інтактному стані та в динаміці вегетативного дисбалансу. По вісі абсцис – час від початку досліду, дні; по вісі ординат – дебіт об’єму, мл/год (M±m, N=15 (інтактний стан), N=5 (дослід)).
було спостерігати закидання дуоденального вмісту в шлунок, про що свідчила наявність жовчних кислот, концентрація яких досягала максимальних величин наприкінці експерименту. При цьому рН шлункового соку змінювався відповідно та на 15-й день дослідження досяг мінімальних значень, що вже не змінювалися до кінця експерименту. Загалом на 7–10-й день у міжтравний період з’явилася кисла шлункова секреція натще, збільшилося виділення слизу й у шлунковому вмісті виявлялися жовчні кислоти. Для перевірки причин появи кисло секреції застосовували атропіновий тест, який був позитивним. У той же час, поява жовчних кислот у шлунковому вмісті обумовлена закиданням дуоденального вмісту в шлунок. Причиною цього дуоденогастрального рефлюксу, у свою чергу, антиперистальтичні скорочення гладеньком’язової мускулатури дванадцятипало кишки, що пов’язане із включенням компенсаторних механізмів, спрямованих на ліквідацію закислення в гастродуоденальній зоні. Перевірка функціональної здатност секреторних залоз шлунка за допомогою гістамінового тесту виявила також зміни резервних можливостей секреторного апарату шлунка (рис. 2, А, Б).
Міоелектрична активність шлунка та дванадцятипалої кишки щурів в інтактному стані. В інтактному стані міоелектрична активність гастродуоденальної зони мала фазний характер, при цьому фаза спокою становила 40-50 хв, фаза нерегулярної піково активності – 50-60 хв, фаза регулярної пікової активності – 5-9 хв. В нтактному стані період основного електричного ритму шлунка склав 11,88±0,20 с (P<0,05), а дванадцятипалої кишки – відповідно 1,61±0,02 с (P<0,05). Однак у ряді експериментів період основного електричного ритму шлунка не перевищував 8,30±0,34 с (P>0,05), а в інших тварин в окремих експериментах цей показник дванадцятипалої кишки досяг 2,10±0,15 с (P<0,05), що вказує на ндивідуальні особливості моторики верхніх відділів шлунково-кишкового тракту.
Міоелектрична активність шлунка та дванадцятипалої кишки, та секреторна діяльність шлунка в інтактних тварин за умов блокування NO-синтази. В інтактних тварин уже через 5–7 хв після введення блокатору NO-синтази в складі міоелектричної активності спостерігалося збільшення амплітуди маскуючих коливань, на тлі яких почали проявлятись поодинокі пікові потенціали, а через 35–45 хв спостерігали електричні коливання стінки шлунка, які були схожі за складом із ІІ-ІІІ фазою міоелектрично активності; така картина зберігалася протягом наступних 75–85 хв. При цьому електричні коливання, які були найбільш подібні до ІІІ фази, спостерігалися в шлунці через 80–100 хв та тривали не більше 10–15 хв. При аналіз міоелектричної активності дванадцятипалої кишки при блокаді нітрергічних механізмів відзначено збільшення амплітуди основного електричного ритму за рахунок підвищення амплітуди першого висхідного коліна повільної хвилі вже через 12–16 хв після введення L-NNA, а через 20 хв спостерігали появу ІІІ фази міоелектричної активності. Треба зазначити, що як у вихідному стані, так після введення L-NNA, міоелектрична активність дванадцятипалої кишки містила нестійкі за появою повільнохвильові коливання, період яких, як правило, наближався за значенням до періоду основного електричного ритму шлунка та становив 6,80±0,15с. Значення основних показників міоелектричної активност шлунка та дванадцятипалої кишки після введення L-NNA на різних етапах розвитку дуоденогастрального рефлюксу наведені в таблиці 1.
Таблиця 1.
Основні показники міоелектричної активності шлунка та дванадцятипалої кишки після введення L-NNA після 3-х, 6-ти, 12-ти днів моделювання дуоденогастрального рефлюксу (M±m)
| Показник | Інтактне (N=12) | Після 3-х днів ДГР (N=9) | Після 6-ти днів ДГР (N=9) | Після 12-ти днів ДГР (N=9) |
| Період ОЕР шлунка | 10,85±0,26* | 11,16±0,39* | 14,66±0,57 | 15,79±0,68 |
| Період ОЕР ДПК | 1,60±0,02 | 1,73±0,03 | 1,96±0,04 | 1,93±0,06** |
| Амплітуда ОЕР шлунка | 40,80±4,08 | 15,12±2,20 | 25,88±3,05 | 13,96±2,52** |
| Амплітуда ОЕР ДПК | 24,12±1,64* | 26,10±1,37 | 54,15±2,97* | 31,28±1,39 |
| Моторний індекс шлунка | 8,25±1,06 | 4,49±1,35 | 7,59±1,03 | 5,99±1,19** |
| Моторний індекс ДПК | 6,68±0,98 | 7,76±1,74 | 13,50±0,67 | 8,88±0,64 |
Примітка: * – P<0,05, **– P<0,01 достовірність різниць за відношенням до відповідних значень вихідного стану; ДГР – дуоденогастральний рефлюкс, ОЕР – основний електричний ритм, ДПК дванадцятипала кишка.
В окремій серії експериментів було вивчено морфофункціональний стан слизової оболонки шлунка інтактних щурів після 12-ти днів щоденного введення L-NNA у прийнятій дозі. У цій серії шлунковий сік збирали протягом однієї години після відповідного збору порції натще. Секреторна діяльність шлунка при цьому характеризувалася різноспрямованими змінами. Так, порції соку натще частіш за все були кислими (рН досягав 0,8–0,9), але наступні порції соку мали значно вищий рН, який становив 3,61±0,60 (N=8). Рівні пепсину в порціях натще рідко були нижчими за 1,50 мг/мл, інколи досягали 1,80 мг/мл, проте концентрація пепсину в базальній порції (загалом за годину) становила 0,69±0,04 мг/мл (N=8), а рівень глікопротеїнів – відповідно 0,045±0,0027 мг/мл (N=8).
Міоелектрична активність шлунка та дванадцятипалої кишки при розвиткові дуоденогастрального рефлюксу. При аналізі міоелектричної активност шлунка та дванадцятипалої кишки в цих експериментах відмічалися значні порушення фазності цього показника, які виражалися як в змінах почерговості фаз, так і в якісному складі останніх. Тому треба підкреслити, що власне фази міоелектрично активності, які спостерігались в цей строк, дещо відрізнялися від так званих «нативних» фаз (які спостерігаються в інтактному стані). Так, домінуючими фазами міоелектричної активності шлунка були пізня ІІ-а фаза, коли спалахи пікових потенціалів спостерігалися практично на кожній 2-й повільній хвилі, та ІІІ-я фаза, коли на тлі основного електричного ритму проявлялася безперервна пікова активність. У вихідному стані міоелектрична активність дванадцятипалої кишки характеризувалася перевагою фази спокою (І-а фаза), яка тривала 60–70 хв та переривалася «активними» періодами – ІІ–ІІІ-я фаза (10–25 хв), коли на тл нерегулярної пікової активності чергувалися відрізки з регулярною піковою активністю (по 10–40 с). При цьому характерними були ознаки аритмії основного електричного ритму шлунка та дванадцятипалої кишки. Значення основних показників міоелектричної активності шлунка та дванадцятипалої кишки на різних етапах розвитку дуоденогастрального рефлюксу наведені в таблиці 2.
Таблиця 2.
Основні показники міоелектричної активності шлунка та дванадцятипалої кишки після 3-х, 6-ти, 12-ти днів моделювання дуоденогастрального рефлюксу (M±m)
| Показник | Інтактне (N=12) | Після 3-х днів ДГР (N=9) | Після 6-ти днів ДГР (N=9) | Після 12-ти днів ДГР (N=9) |
| Період ОЕР шлунка | 11,88±0,19 | 13,00±0,44* | 13,75±0,25* | 13,56±0,73* |
| Період ОЕР ДПК | 1,61±0,03 | 1,73±0,03** | 1,90±0,04** | 1,77±0,03** |
| Амплітуда ОЕР шлунка | 40,41±3,22 | 17,79±2,86** | 22,92±2,45** | 47,05±5,63 |
| Амплітуда ОЕР ДПК | 20,63±0,96 | 25,90±1,42* | 44,94±2,01** | 29,79±1,44* |
| Моторний ндекс шлунка | 6,77±0,85 | 5,27±1,49 | 5,87±1,23 | 12,30±1,72** |
| Моторний ндекс ДПК | 4,81±0,73 | 6,69±1,33 | 11,31±1,03** | 8,06±1,34** |
Примітка: * – P<0,05, **– P<0,01 достовірність різниць за відношенням до відповідних значень інтактного стану. Позначення такі ж, як в табл. 1.
Після 6 днів моделювання у складі міоелектричної активності шлунка з’явилися неперіодичн високоамплітудні коливання, а сама картина цього показника гастродуоденально зони більшою мірою відповідала початку II нативної фази. Треба зазначити, що основні електричні ритми шлунка та дванадцятипалої кишки не завжди чітко виявлялися протягом експерименту. Найбільш характерною зміною міоелектрично активності була поява періодичної пікової активності дванадцятипалої кишки. Частота пікових спалахів склала 2,76 ± 0,07 кол/хв, а амплітуда досягала 60,0 мкВ вище. На 12-й день експерименту зберігалася перевага II фази з вираженою нерегулярної піковою активністю й більш «збудженим» станом міоелектрично активності. Останнє було більш характерним для дванадцятипалої кишки, ніж для шлунка. У багатьох експериментах спостерігалася тенденція до поширення ритмів дванадцятипалої кишки на шлунок, особливо тоді, коли реєструвалась I фаза міоелектричної активності. При цьому на електрогастроміограмі можна було спостерігати періодичні високоамплітудні коливання з періодом 83,07±0,29 с амплітудою 10,56±0,06 мкВ, а на електродуоденоміограмі їм у часі відповідали коливання амплітудою 11,76±0,17 мкВ. Окрім змін фазної природи міоелектрично активності в цей строк спостерігалися також зміни періоду та амплітуди основного електричного ритму. Так, період основного електричного ритму шлунка після 12-ти днів дослідження склав (за даними двохгодинного спостереження) 13,56±0,73 с (N=9), що фактично не відрізнялось від значень 3-го та 6-го дня, однак амплітуда основного електричного ритму шлунка досягла 47,05±5,63 мкВ (N=9), що становило двократне збільшення за відношенням до показника 6-го дня та практично відповідало значенню амплітуди основного електричного ритму нтактного шлунка. Період основного електричного ритму дванадцятипалої кишки помітно знизився (на 6,84%, P<0,05) у порівнянні із значенням 6-го дня та становив 1,77±0,03 с (N=9), і отже суттєво не відрізнявся від показника 3-го дня. Також відбулося зниження амплітуди основного електричного ритму дванадцятипалої кишки на 33,71% (P<0,05) за відношенням до значення 6-го дня, яка (амплітуда) несуттєво перевищувала показник 3-го дня та склала 29,79±1,44 мкВ (N=9). Отже, при моделюванні дуоденогастрального рефлюксу періоди основного електричного ритму шлунка та дванадцятипалої кишки поступово збільшувалися до 6-го дня, а на 12-й день спостерігалося зниження цього показника дванадцятипалої кишки та відповідно тенденція до зниження показника в шлунці.
Міоелектрична активність шлунка та дванадцятипало кишки за умов блокування NO-синтази на тлі моделювання дуоденогастрального рефлюксу. Після 3-х днів моделювання дуоденогастрального рефлюксу не спостерігалися будь-які достовірні відмінност міоелектричної активності шлунка: протягом часу спостереження переважала фаза нерегулярної пікової активності, яка або «посилювалася» (збільшення амплітуди та кількості нетипових коливань та пікових потенціалів), або ж «послаблювалася» неперіодичним чином, при цьому такі явища не завжди були пов’язані із введенням блокатору NO-синтази. У більшості експериментів цієї серії після введення блокатору NO-синтази відмічалося «посилення» вже зафіксованих явищ міоелектрично активності, які були описані для вихідного стану, а також поява кількісних різниць у їх виявленні. Після 3-х днів моделювання дуоденогастрального рефлюксу через 20 хв після введення L-NNA спостерігалося вкорочення періоду базального ритму шлунка (Р<0,05, табл. 2). У той же час, такі умови не викликали будь-яких достовірних змін з боку періоду базального ритму дванадцятипало кишки, зміни якого у вихідному стані мали вигляд періодичних з «періодом» близько 2-х годин. При блокуванні NO-синтази після 6-ти днів моделювання дуоденогастрального рефлюксу зміни міоелектричної активності шлунка та дванадцятипалої кишки не були узгоджені в часі. Першою на введення L-NNA відреагувала дванадцятипала кишка: через 1,5–2,0 хв після ін’єкції блокатору спостерігалося проявлення регулярної пікової активності (амплітудою 20–40 мкВ), при цьому пікова активність «розташовувалася» у вигляді відрізків по 21,09±1,76 с (N=8). На відміну від дванадцятипалої кишки, помітні зміни міоелектрично активності шлунка спостерігалися лише після 30–35-ти хв та полягали в наявност нерегулярної пікової активності (яка тривала 35–45 хв) та більш вираженій аритмії базального ритму. Окрім того, в цей час стало помітним нестабільне проявлення періодичних повільних коливань на тлі основного електричного ритму шлунка, які за своїм періодом практично відповідали значенням базального ритму дванадцятипалої кишки; таке явище ми охарактеризували як ретроградне розповсюдження основного електричного ритму останньої на ділянку пілоричного відділу шлунка.
Після 12-ти днів блокада нітрергічної ланки регуляц обумовила збільшення загальної тривалості «активних» фаз міоелектрично активності шлунка (ІІ–ІІІ-я фази). Перші ознаки цього були помітні вже через 5–10 хв після ін’єкції, коли в складі електрогастроміограми з’явилися періодичні атипові коливання, які мали період 18,76±0,87 с, амплітуду 142,00±9,01 мкВ (N=9) та тривали 10–12 хв. При аналізі змін міоелектрично активності дванадцятипалої кишки після введення L-NNA передусім треба зазначити наявність безперервної регулярної пікової активності, яка починалася через 2–5 хв після ін’єкції блокатору та тривала протягом 2-х годин. При цьому спалахи пікових потенціалів розташовувалися або на кожній 2-й хвилі основного електричного ритму, або ж на відрізках повільних коливань, що чергувалися, та містили пікову активність на кожній 2-й/4-й/6-й/8-й хвилі.
За відношенням до відповідного вихідного стану (табл. 1) після введення L-NNA період базального ритму шлунка в контрольній груп (інтактний стан) знизився на 8,67% (Р<0,05, табл. 2), після 3-х днів моделювання дуоденогастрального рефлюксу – знизився на 14,15% (Р<0,05), після 6-ти днів – збільшився на 6,62% (Р>0,05), а після 12-ти днів відбулося його підвищення на 16,45% (Р<0,05). На відміну від періоду базального ритму шлунка, цей показник дванадцятипалої кишки після введення L-NNA за відношенням до відповідного вихідного стану достовірно не змінився ні в інтактному стані, ні після 3-х днів дослідження, ні після 6-ти днів, але після 12-ти днів виявив збільшення на 9,04% (Р<0,05).
Діяльність секреторних залоз шлунка при розвитков дуоденогастрального рефлюксу. Після 3-х днів дослідження спостерігали помітне збільшення концентрації пепсину (на 18,10%, Р>0,05), значні підвищення концентрац глікопротеїнів (на 41,18%, Р<0,05), рівня рН (на 32,61%, Р<0,05) при, однак, недостовірному збільшенні об’єму шлункового аспірату (рис. 3, А, Б).
Як видно з рисунку 3 (А), після 6-ти днів збільшився рівень рН шлункового соку (на 62,20% за відношенням до вихідного стану, Р<0,05) при збереженні об’єму шлункового соку, характерного для 3-го дня дослідження та поверненні рівней пепсину та глікопротеїнів до значень інтактного стану. При цьому після 12-ти днів у шлунковому соці спостерігали зниження концентрац пепсину (на 36,65%, Р<0,05), збільшення об’єму шлункового соку (на 85,35%, Р<0,05) та підвищення концентрації глікопротеїнів (рис. 3, Б) у 5,88 разів (Р<0,001) за відношенням до інтактного стану. Це супроводжувалося підвищенням кислотності містива шлунка, що виражалося у зниженні рН шлункового соку до показників інтактного стану.
Діяльність секреторних залоз шлунка за умов блокування нітрергічних механізмів на тлі розвитку дуоденогастрального рефлюксу. Блокування NО-синтази у тварин контрольної групи обумовило зсуви в характері шлункової секреції, які виражалися як у збільшенн об’єму шлункового соку (на 20,25%, Р>0,05) та рівня глікопротеїнів (на 79,41%, Р<0,05), так і в зниженні рівня пепсину (на 19,46%) (P<0,05) та значення рН (на 17,13%, P<0,05). Після 3-х днів моделювання дуоденогастрального рефлюксу введення блокатору зумовило підвищення концентрації глікопротеїнів (майже в 2,20 рази, Р<0,05), об’єму шлункового соку (на 85,71%, Р>0,05) та кислотності останнього, що виразилося у збільшенні рН соку на 82,65% (Р<0,05). Блокування NO-синтази після 6-ти днів дослідження, навпаки, зумовило зниження рівня пепсину (на 26,63%, Р>0,05) в шлунковому соці та значне підвищення рівня рН (майже в 2 рази, Р<0,05) за відношенням до вихідного стану при незначних змінах об’єму шлункового аспірату та незмінності концентрації глікопротеїнів в шлунковому соці. На 12-й день при блокаді нітрергічних механізмів спостерігалося зменшення об’єму шлункового соку (на 41,46%, Р<0,05) по відношенню до вихідного стану при практично незмінних рівнях пепсину, глікопротеїнів та кислотності соку.
Узагальнення результатів дослідження. При моделюванні порушень морфофункціональної цілісності гастродуоденальної зони фази міоелектричної активності шлунка та дванадцятипалої кишки відрізнялися від відповідних фаз інтактної міоелектричної активності. До цих змін міоелектрично активності належать особливості пікової активності та загальний характер проявлення періодичних електричних коливань стінки органа, при цьому була порушена почерговість фаз. Тому більш доцільно казати не про періодичне проявлення окремих фаз як складових частин мігруючого моторного комплексу, а про часове домінування самих фаз або їх комплексів у цілісному патерн міоелектричної активності: фази відносного спокою (І-а – початкова ІІ-а фаза) та «активні» фази (пізня ІІ-а – ІІІ-я фаза). На початкових етапах моделювання вегетативного дисбалансу спостерігалася тенденція до зрівняння тривалостей «активних» фаз (II-III-фази) і фази спокою (I фаза) міоелектричної активност шлунка. Результати порівнянь змін моторики шлунка та його секреції вказують на більшу чутливість першої до змін балансу симпато-парасимпатичних впливів у регуляторній системі, ніж діяльність його секреторних залоз. При цьому ретроградне поширення основного електричного ритму шлунка та дуоденогастральний рефлюкс, які мали місце на 7–10 день, є адаптаційно-компенсаторною реакцією місцевої нейроімуноендокринної системи на закислення шлункового вмісту. Переломним” періодом в порушенні регуляції моторно-секреторної діяльност шлунка є період 10–15-го днів, про що свідчить помітне зниження впливу симпатоадреналової системи. На наш погляд, у реалізації даних ефектів у цей період більше значення набувають неадренергічні нехолінергічні механізми, значна роль яких у регуляції моторики шлунково-кишкового тракту підтверджується рядом авторів [Boeckxstaens et al., 1992; D’amato et al., 1992; Meulemans et al., 1995], а деякі дослідники ставлять вказані механізми за регуляторною значимістю на один рівень з холінергічною регуляцією [Galligan et al., 1986]. Наприкінці експерименту (30–35-й день) спостерігали зниження діяльност секреторних залоз шлунка при стимуляції гістаміном, що можна пояснити явищем «відмови в регуляції», якому одночасно сприяли великі зміни моторики шлунка, що загалом можна розглядати як прояв декомпенсаційних перебудов регуляторних механізмів. Детальна участь вказаних механізмів у динаміці місцевих регулятивно-морфофункціональних зсувів з’ясовувалася нами в серіях експериментів на щурах.
На всіх етапах моделювання дуоденогастрального рефлюксу спостерігалося домінування активних фаз міоелектричної активності шлунка та дванадцятипалої кишки. З урахуванням точок зору ряду авторів [Takeuchi et al., 1997, 2000; Tanaka et al., 2001], можна припустити, що це пов’язано з: 1) безпосереднім впливом жовчі як ушкоджуючого фактору слизової оболонки шлунка; та з 2) порушеннями нейроімуноендокринно регуляції періодичної діяльності гастродуоденальної зони під впливом несприятливих факторів. Так, при дії таурохолатів авторами [Takeuchi et al., 2000] було встановлено збільшення рівней порожнинних простагландинів, що вказувало на активацію захисних механізмів слизової оболонки шлунка. При цьому простагландини відомі своїм гальмівним впливом у відношенні кислої шлунково секреції [Khattab et al., 2001], зміни якої натще у здорових людей оборотно пов’язані із моторною активністю шлунка [McCallum, 1997].
Результати даної серії наших експериментів частково узгоджуються із даними інших авторів [Вильгельм и др., 1994; Маев и др., 2000], які вказують на часте поєднання дуоденогастрального рефлюксу у людини із зниженням моторної активності дванадцятипалої кишки. У наших дослідженнях у переважній кількості спостережень 3-го дня та в ряді експериментів 6-го та 12-го днів у складі міоелектричної активності дійсно переважали фази відносного спокою (І – початкова ІІ-а фаза), на тлі яких час від часу виникали пікові або атипові коливання. При цьому відзначалася тенденція до протилежного розповсюдження дуоденального ритму – на ділянку антрального відділу шлунка, що свідчило про можливість закиду дуоденального вмісту в шлунок. Дійсно, після 6-ти та 12-ти днів в шлунковому соці виявлялися жовчні кислоти.
При обговоренні змін моторики при дуоденогастральному рефлюксі не можна не враховувати викид серотоніну з ЕС-клітин у кровотік, який має місце при стресі, стимуляції вегетативно нервової системи [Ahlman et al., 1984] та впливає на моторно-секреторну діяльність шлунка, та стан його слизової оболонки, що є, в основному, результатом вазоконстрикторної дії аміну [Amemiya et al., 1996; Takada et al., 1999]. Надлишок серотоніну, в свою чергу, зумовлює шлункове виразкоутворення та дуоденогастральну дискінезію [Шемеровский, 1989; 1998], які й спостерігалися в наших експериментах. На відміну від інтактного стану, після 3-х та 6-ти днів дослідження тонус (за даними амплітудної характеристики міоелектрично активності) дванадцятипалої кишки значно перевищував тонус антрального відділу шлунка, що могло бути однією з причин закиду дуоденального вмісту в шлунок. І лише після 12-ти днів дослідження відбулося різке падіння амплітуди основного електричного ритму дванадцятипалої кишки при одночасному зростанні цього показника в шлунці, що можна пояснити активацією місцевих адаптаційно-компенсаторних механізмів, спрямованих на нормалізацію антероградного розповсюдження моторних ритмів гастродуоденальної зони.
Результати наших досліджень свідчать на користь того, що нітрергічні механізми мають менше значення для регуляц моторики дванадцятипалої кишки, ніж моторики шлунка. На 6–12-й день відбувалася протилежна зміна ролі нітрергічних механізмів, коли NO почав відігравати роль стимулятора гладеньких м’язів пейсмекерних зон антрального відділу шлунка, що, можливо, є проявом адаптаційних механізмів, спрямованих на збільшення частоти основного електричного ритму шлунка і як наслідок – тонусу та прискорення видалення екзогенного подразника (жовчі). Наші результати вказують на те, що в нтактному стані NO дійсно здатний справляти гальмівний вплив на парієтальн клітини слизової оболонки шлунка, при цьому блокада NO-синтаз супроводжувалася також зниженням рівня пепсину та значним збільшенням концентрац глікопротеїнів. Частково такі ефекти можна пояснити тим, що NO в інтактному стані має стимулюючий вплив у відношенні циклооксигенази, в результаті чого збільшується синтез простагландинів [Khattab et al., 2001], які, як відомо, здатні гальмувати кислу шлункову секрецію. Тенденція до зниження рівня пепсину та підвищення концентрації глікопротеїнів під дією L-NNA, які спостерігалися в наших дослідженнях, вказують на можливу стимулюючу дію NO у відношенн діяльності головних клітин та гальмівну – у відношенні поверхнево-епітеліальних клітин. Такий вплив NO, на нашу думку, має бути опосередкованим, адже в доступній літературі ми не знайшли будь-яких відомостей щодо наявност специфічних клітинних утворень, які б зумовлювали дію NO на ці клітини слизово оболонки шлунка.
На 12-й день нашого дослідження перебудови місцевих регуляторних механізмів шлункової секреції виразилися у зменшенн обсягу шлункового соку за умов блокади NO-синтази при практично незмінених концентраціях пепсину, глікопротеїнів, рівня рН. З урахуванням відомих досліджень [Takeuchi et al., 1995, 2000], такі факти вказують на вірогідну нверсію впливу нітрергічної ланки в регуляції кислої шлункової секреції, коли NO починає грати роль стимулятора останньої.
ВИСНОВКИ
1. При вегетативному дисбалансі та дуоденогастральному рефлюксі відбуваються різноспрямовані перебудови місцевих нейроендокринних регуляторних механізмів періодичної моторно-секреторної діяльності гастродуоденальної зони, що супроводжується адаптаційними та патологічними змінами її слизової оболонки, та декомпенсаційними змінами різних ланок регуляції.
2. Зміни моторно-секреторних ритмів шлунка при розбалансуванні місцевих регуляторних механізмів протікали в три стадії: на першій стадії спостерігалося зрівняння тривалостей періодів «збудження» (II-а та III-я фази міоелектричної активності) з періодами спокою (I-а фаза міоелектричної активності) при одночасному посиленні активност поверхнево-епітеліальних та зниженні активності парієтальних клітин слизово оболонки шлунка; на другій стадії (7–15-й день) десинхронізація основного електричного ритму шлунка супроводжувалася підвищенням функціонально активності секреторних залоз шлунка; на третій стадії разом із поступовим збільшенням тривалості періодів спокою, амплітуди первинної хвил міоелектричної активності, періоду основного електричного ритму шлунка та зменшенням сумарної тривалості «активних» фаз міоелектричної активност відбувалося стрімке зниження функціональної здатності секреторних залоз шлунка та закидання дуоденального вмісту в шлунок.
3. До 6-го дня розвитку дуоденогастрального рефлюксу відбувалася поступова адаптація регуляторних систем слизової оболонки шлунка до дії несприятливих факторів, що проявлялося у зниженні «агресивних» властивостей шлункового соку, появі періодичних високоамплітудних атипових коливань на електродуоденоміограм та дестабілізації основних електричних ритмів шлунка та дванадцятипалої кишки з поступовим зниженням їх частоти, а також перевагою ІІ фази міоелектрично активності.
4. В інтактному стані у щурів нітрергічні механізми регуляції мають гальмівний вплив у відношенні парієтальних та поверхнево-епітеліальних клітин слизово оболонки шлунка, а також скоротливої активності шлунка та дванадцятипало кишки. При розвитку дуоденогастрального рефлюксу NO-ергічна регуляція зберігає свій гальмівний вплив у відношенні скоротливої активност гастродуоденальної зони шляхом пригнічення міогенної пікової активност антрального відділу шлунка та цибулини дванадцятипалої кишки.
5. При розвиткові дуоденогастрального рефлюксу відбувається перебудова місцево нейроендокринної регуляції гастродуоденальної зони, однією зі шляхів якої може виступати протилежна зміна (інверсія) впливу NO-ергічно ланки у напрямку діяльності парієтальних клітин шлунка та її вибіркова протилежна зміна впливу на періодичну міогенну активність шлунка та дванадцятипалої кишки.
6. Зміна ролі нітрергічної ланки регуляції у відношенні моторики гастродуоденальної зони при розвитку дуоденогастрального рефлюксу протікала в дві стадії: на першій стадії (6-й день дослідження) зміни торкнулися здебільшого механізмів генерації основного електричного ритму шлунка та в меншій мірі – основного електричного ритму дванадцятипалої кишки, на другій стад (12-й день дослідження) – відбулися зміни у відношенні діяльності секреторного апарату шлунка.
7. Результатом адаптаційно-компенсаторних перебудов регулятивно-ефекторної системи періодичної діяльності гастродуоденальної зони при розвитку дуоденогастрального рефлюксу є підвищення активності парієтальних та поверхнево-епітеліальних клітин слизової оболонки шлунка, пригнічення діяльності її головних клітин, а також зміна ролі нітрергічної ланки регуляції в напрямку стимуляції частоти основних електричних ритмів гастродуоденальної зони та кислотосекреторного апарату шлунка.
СПИСОК РОБІТ, ОПУБЛІКОВАНИХ ЗА ТЕМОЮ ДИСЕРТАЦІЇ
1. Руденко А. І., Трушенко О. С., Мурзін О. Б. Періодична діяльність секреторних залоз шлунка при виразковому ушкодженн гастродуоденальної зони // Вісник Дніпропетровського університету. Серія: Біологія. Екологія. – 2006. – Вип. 14. – Т. 1. – С. 154–159. (Особистий внесок здобувача: участь у проведенні експериментів, оформлення отриманих результатів у вигляді статті).
2. Трушенко О. С., Мурзін О. Б., Мосійчук Л. М., Руденко А. І. Нітрергічні механізми регуляції діяльності секреторних залоз шлунка при дуоденогастральному рефлюксі у щурів // Вісник проблем біології і мед. – 2007. – № 3. – С. 73–78. (Особистий внесок здобувача: проведення експериментів, оформлення одержаних результатів у вигляді статті).
3. Руденко А. І., Трушенко О. С. Моторна діяльність гастродуоденальної зони при дуоденогастральному рефлюксі у щурів // Гастроентерологія / Міжвід. збірник. – Вип. 38. – Д., 2007. – С. 119–127. (Особистий внесок здобувача: проведення експериментів, аналіз та систематизація одержаних результатів, та оформлення їх у вигляді статті).
4. Трушенко О. С., Мурзін О. Б., Руденко А. І. Періодична моторно-секреторна діяльність шлунка при дисбалансі його регуляторних механізмів // Фізіологічний журнал. – 2007. – № 5. – С. 91–98. (Особистий внесок здобувача: участь у проведенні експериментів, виконання статистично обробки даних, узагальнення результатів роботи та оформлення їх у вигляд статті).
5. Trushenko A.S., Murzin A.B., Rudenko A.I. Peculiarities of the stomach and duodenal myoelectrical activity in different gastro-duodenal injuries / II International conference Neural-humoral and cellular regulatory mechanisms of digestion processes (October 5-7, 2005, Kyiv, Ukraine). – Kyiv, 2005. – P. 34.
6. Трушенко О.С., Мурзін О.Б., Руденко А.І. Періодичні ритми гастродуоденальної зони при порушенні співвідношень вегетативних впливів // Матер. Всеукр. наук.-практ. конф. «Вклад молодих вчених в розвиток медичної науки і практики» (12 квітня 2006 р.). – Харків, 2006. – С. 125.
7. Трушенко А.С., Мурзин А.Б., Руденко А.И. Ультрадианные ритмы гастродуоденальной зоны при повышении активности парасимпатического отдела вегетативной нервной системы //Збірник тез VIII Міжнар. наук.-практ. конф. «Людина і космос. Ракетно-космічна техніка» (13-14 квітня 2006р). – Дніпропетровськ, 2006. – С. 212.
8. Трушенко О. С., Мурзін О. Б., Руденко А. І. Міоелектрична активність шлунка та дванадцятипалої кишки при ушкодженн гастродуоденальної ділянки. Матеріали 17-го з’їзду Укр. фізіол. тов. з міжнар. участю. Чернівці, 18-20 травня, 2006 // Фізіологічний журнал. – 2006. – Т. 52, 2. – С. 169.
9. Трушенко О. С., Мурзін О. Б., Руденко А. І. Адаптаційно-компенсаторні перебудови мігруючого міоелектричного комплексу та секреторної діяльності гастродуоденальної зони в умовах дисбалансу регуляторних механізмів // Матер. Міжнар. наук. конф. «Механізми функціонування фізіологічних систем» (8–11 листопада 2006 р.). – Львів, 2006. – 143–144.
10. Трушенко О. С., Мурзін О. Б., Руденко А. І. Періодичні моторно-секреторні ритми гастродуоденальної зони в динаміц дуоденогастрального рефлюкса у щурів // Матер. Всеукр. наук. конф. «Сучасн питання фізіології та медицини», присв. 85-річчю заснування каф. фізіолог людини і тварин та 100-річчю з дня народження П. Є. Моцного (26–28 вересня 2007 р). – Д., 2007. – С. 91.
АНОТАЦІЇ
Трушенко О.С. Координація періодичної діяльност гастродуоденальної зони при порушенні її функціонально-морфологічно цілісності. – Рукопис.
Дисертація на здобуття наукового ступеня кандидата біологічних наук за фахом 13.00.13 – фізіологія людини і тварин. – Таврійський національний університет імені В.І. Вернадського, Симферополь, 2008.
Вивчена моторно-секреторна діяльність шлунка та дванадцятипалої кишки при порушеннях функціонально-морфологічної цілісност гастродуоденальної зони у собак та щурів. Встановлено, що вегетативний дисбаланс обумовив зміни тривалості фаз періодичної моторно-секреторно діяльності шлунка, появу кислої секреції натще, посилення відповідної реакц шлункових залоз на екзогенну стимуляцію гістаміном та явище „відмови в регуляції”, при цьому моторні ритми виявились більш чутливими до регуляторних зсувів, ніж секреторна діяльність шлунка. Моделювання дуоденогастрального рефлюксу у щурів зумовило порушення фазності міоелектричної активност гастродуоденальної зони, що супроводжувалося переважанням пікової активності та тенденцією до розповсюдження дуоденальних ритмів на шлунок. Це супроводжувалося зниженням агресивних властивостей шлункового соку (6 день) та збільшенням його об’єму (12-й день). При цьому NO-ергічна регуляція зберегла свій гальмівний вплив у відношенні скоротливої активності антрального відділу шлунка та цибулини дванадцятипалої кишки, а на 12-й день відбулося зниження обсягу шлункового соку при блокаді NO-синтаз при практично незмінних концентраціях пепсину, глікопротеїнів та рН.
Ключові слова: вегетативний дисбаланс, дуоденогастральний рефлюкс, гастродуоденальна зона, міоелектрична активність, шлункова секреція, NO-синтаза
Трушенко А.С. Координация периодической деятельности гастродуоденальной зоны при нарушении ее функционально-морфологической целостности. – Рукопись.
Диссертация на соискание ученой степени кандидата биологических наук по специальности 03.00.13. – физиология человека и животных. Таврический национальный университет имени В.И. Вернадского, Симферополь, 2008.
В работе выявлены основные закономерности динамики моторно-секреторной деятельности желудка и двенадцатиперстной кишки при моделировании вегетативного дисбаланса у собак и при развитии дуоденогастрального рефлюкса у крыс. Изучена также роль нитрэргического звена регуляции в координации моторно-секреторных паттернов гастродуоденальной зоны при моделировании дуоденогастрального рефлюкса у крыс.
Установлено, что при моделировании вегетативного дисбаланса у собак происходят изменения функционально-морфологической целостности слизистой оболочки желудка и его моторной активности. При этом изменения моторно-секреторных ритмов желудка протекали в 3 стадии. На первой стадии (до 7-го дня) наблюдалось уравнивание длительностей «активных» периодов миоэлектрической активности с периодами покоя при одновременном увеличении активности поверхностно-эпителиальных и снижении активности париетальных клеток слизистой оболочки желудка. На второй стадии (7–15-й день) десинхронизация основного электрического ритма желудка сопровождалась повышением функциональной активности секреторных желез желудка. На третьей стадии вместе с постепенным увеличением длительности периодов покоя, амплитуды первичной волны миоэлектрической активности, периода основного электрического ритма желудка и уменьшением суммарной длительности «активных» фаз миоэлектрической активности наблюдалось резкое снижение функциональных возможностей секреторных желез желудка и дуоденогастральный рефлюкс.
При развитии дуоденогастрального рефлюкса до 6-го дня наблюдалась адаптация регуляторных систем слизистой оболочки желудка к действию неблагоприятных факторов, что проявлялось в снижении «агрессивных» свойств желудочного сока, появлению периодических атипичных колебаний на электродуоденомиограмме, и дестабилизации основных электрических ритмов желудка и двенадцатиперстной кишки с постепенным снижением их частоты, а также преобладанием ІІ фазы миоэлектрической активности. Это свидетельствовало о возможности заброса дуоденального содержимого в желудок, что и подтверждалось наличием желчных кислот в желудочном соке с 6 дня эксперимента. В конце эксперимента (12 день) резко падает амплитуда основного электрического ритма двенадцатиперстной кишки при одновременном повышении этого показателя в желудке, что указывает на активацию местных адаптационно-компенсаторных механизмов, направленных на нормализацию антероградного распространения моторных ритмов гастродуоденальной зоны. Указанные изменения моторики сопровождались увеличением активности париетальных и поверхностно-эпителиальных клеток слизистой оболочки желудка, а также угнетением деятельности ее главных клеток. Установлено, что при развитии дуоденогастрального рефлюкса NO-эргическая регуляция сохраняет свое тормозное влияние в отношении сократительной активности гастродуоденальной зоны путем угнетения миогенной пиковой активности антрального отдела желудка и луковицы двенадцатиперстной кишки. Наблюдавшиеся перестройки местной нейроэндокринной регуляции одним из механизмов могли иметь противоположное изменение (инверсия) влияния NO-эргического звена в направлении деятельности париетальных клеток желудка и его выборочное противоположное изменение влияния на периодическую миогенную активность желудка и двенадцатиперстной кишки. При этом изменение роли NO-эргического звена регуляции в отношении моторики гастродуоденальной зоны коснулось первично (6 день) и в большей мере механизмов генерации базального ритма желудка, в меньшей степени – базального ритма двенадцатиперстной кишки, и вторично (12 день) – активности секреторного аппарата желудка.
Ключевые слова: вегетативный дисбаланс, дуоденогастральный рефлюкс, гастродуоденальная зона, миоэлектрическая активность, желудочная секреция, NO-синтаза
Trushenko A. S. «Coordination of the periodic activity of the gastroduodenal zone in its function-and-morphological integrity disturbance». – Manuscript.
Dissertation for the candidate of biological sciences degree by specialty 03.00.13. – human and animal physiology. – V. I. Vernadskiy Taurida National University, 2008.
The motor-and-secretor activity of the stomach and duodenum in disturbances of the functional-and-morphological integrity of gastroduodenal zone was researched in dogs and rats. It was found out that vegetative system disbalance had caused changes of acid secretion in the fasted state, increase of gastric secretor response to exogenous histamine stimulation and a case of “regulatory rejection”, and motor rhythms appeared more sensitive for the regulatory changes than secretor activity of the stomach. In rats duodenogastric reflux modelling caused disturbances of a phasic nature of the gastroduodenal myoelectrical activity, which was associated with the spike-activity predominance and the tendency of the duodenal rhythms to diffuse on the stomach. It was associated with the decrease of the aggressive properties of gastric juice (6 day) and the increase of its volume (12 day). It was found out that the NO-ergic regulation had preserved its inhibitory influence on the contractile activity of the antrum and duodenal bulb, and the decrease of the gastric juice volume could be fixed in NO-syntase-blocked state with nearly unchanged pepsin and glycoprotein concentrations, and pH level.
Keywords: vegetative disbalance, duodenogastric reflux, gastroduodenal zone, myoelectrical activity, gastric secretion, NO-syntase.
Підписано до друку 3.04.2008. Формат 60х90/16. Папір друкарський. Друк плоский.
Гарнітура Times New Roman. Умовн. друк. арк. 1,0. Тираж – 100 прим. Замовлення №
Друкарня ДНУ, 49050, м. Дніпропетровськ, вул. Наукова, 5.
© 2009 База Рефератов