
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Молекулярные механизмы передачи импульса в мембранах нейронов. Ионные каналы, рецепторы
Реферат: Молекулярные механизмы передачи импульса в мембранах нейронов. Ионные каналы, рецепторы
Молекулярные механизмы передачи импульса в мембранах нейронов. Ионные каналы, рецепторы
1. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В НЕРВНЫХ КЛЕТКАХ
На каждом этапе рассмотрения биохимии нервной системы приходится вновь обращать внимание читателя на то обстоятельство, что нейроны способны выполнять свои функции только благодаря особым свойствам их наружной мембраны. Мембрана нейрона имеет специальные молекулярные устройства, которые позволяют ей генерировать, проводить и воспринимать нервный импульс, практически мгновенно изменять ионную проницаемость и создавать за счет этого трансмембранный ионный ток. Этот комплекс молекулярных событий приводит к направленному распространению нервного импульса по аксону на очень большие расстояния.

Способность к проведению нервного импульса в аксонах обусловлена, с одной стороны, наличием в его мембранах специфических белковых комплексов, которые представляют собой ионные каналы, управляемые электрическими потенциалами, с другой стороны, наличием белковых структур, поддерживающих ионные градиенты в мембранах, — так называемых ионных насосов.
Насосы расходуют метаболическую энергию для перемещения ионов против концентрационных градиентов между вне- и внутриклеточной средой. Особенно важны различия в концентрациях ионов Na, К и Са. Наружная среда приблизительно в десять раз богаче ионами Na, чем внутренняя > а внутренняя среда в десятки раз богаче ионами К, чем наружная. Внеклеточные концентрации Са+ в сотни-тысячи раз выше внутриклеточных.
Ионы Na и К могут медленно проникать через поры в клеточной мембране по градиенту, поэтому ионные насосы непрерывно производят обмен вошедших в клетку ионов натрия на ионы калия из внешней среды, такое откачивание ионов натрия осуществляется внутренним мембранным белком — Na+, К+-АТФазой или Na-насосом. Существуют и другие типы ионных насосов, преимущественно называемых по типу ионов, которые они транспортируют, например Са-насосы, К-насосы и т.д..

Модель генерации нервного импульса, созданная А. Ходжкиным и А. Хаксли применительно к аксону, описывает проведение электрического сигнала путем изменения проницаемости для ионов натрия и калия. Эта модель, ставшая классической, принесла авторам известность и Нобелевскую премию в 1956 г. Основная идея модели генерации нервного импульса сводится к следующему: механизмы ионной проницаемости натрия и калия работают независимо друг от друга и описываются с помощью констант скоростей реакции, зависящих от единственной переменной — мембранного потенциала. С помощью экспериментальных подходов эта теоретическая модель была успешно подтверждена.
Поскольку концентрация ионов натрия и калия по ту и другую сторону мембраны различаются, внутренняя область аксона имеет значительный отрицательный потенциал по отношению к наружной среде. Когда нервный импульс возникает в основании аксона, трансмембранная разность потенциалов в этом месте локально понижается. Это ведет к тому, что непосредственно за этой зоной с измененным потенциалом вдоль аксона открываются ионные каналы для входа ионов Na. Процесс является самоусиливающимся: поток ионов натрия через мембрану приводит к открыванию все большего числа ионных каналов. Затем натриевые каналы закрываются, но вслед за этим открывается другая группа каналов — для ионов К, которые выходят наружу. Этот поток восстанавливает потенциал внутри аксона до потенциала покоя. Резкий скачок потенциала или электрический "спайк" называется потенциалом действия и является электрическим выражением нервного импульса.
Итак, возникновение быстрых импульсных сигналов связано с работой ионных каналов. Ионные каналы — это макромо-лекулярные комплексы, которые образуют сквозные гидрофильные поры в липидном матриксе и способны регулировать транспорт ионов через мембрану клетки. Другими словами, ионные каналы представляют собой ионселективный фильтр, способный избирательно регулировать проницаемость клетки для ионов. Так, работа одного ионного канала способна изменять ионные токи от 2 до 10 рА, что соответствует транспорту от 12 до 60x10 моновалентных катионов в секунду. Такая величина обменного процесса ионов в клетке превосходит во много раз известные до сих пор ферментные или транспортные механизмы и хорошо согласуется с теоретическими расчетами, сделанными для модельной поры.
Ионные каналы имеют два фундаментальных свойства: они способны избирательно пропускать ионы и имеют механизм контроля за скоростью перемещения ионов — воротные токи. Однако избирательность каналов для определенных ионов не является абсолютной, так как они могут в определенной степени пропускать и "чужие" ионы, сходные по заряду или размерам.
Механизм селективности ионных каналов определяется взаимодействием между ионами и специфическим структурным участком канала, его воротами. Воротные механизмы, регулирующие открывание и закрывание мембранных каналов, представлены двумя типами. Существуют каналы, которые открываются и закрываются в ответ на изменения потенциалов, т.е. управляются электрически. Второй тип воротного механизма связан с работой ионных каналов, открываемых в ответ на химический сигнал, т.е. управляемых химически.
Деполяризация, связанная с потенциалом действия, распространяется вдоль аксона как волна электрической активности. Главное преимущество электрического проведения импульса по аксону состоит в том, что возбуждение быстро распространяется на большие расстояния без какого-либо ослабления сигнала. Для возникновения серии нервных импульсов необходимо сложное взаимодействие разных ионных каналов, включая электроуправляемые и хемоуправляемые ионные каналы. Все нервные импульсы имеют практически одинаковую амплитуду; кодирование информации на этом уровне происходит за счет разной частоты, генерируемой в единицу времени. В общем, чем сильнее сигнал, тем выше частота разрядов.
2. Na-КАНАЛЫ
Потенциал-зависимые Na-каналы — обязательный элемент внешней мембраны нейронов. В последние годы благодаря обнаружению специфических блокаторов электровозбудимых натриевых каналов удалось раскрыть молекулярную структуру каналов и, в частности, выделить составляющий их белок в индивидуальном виде.
Одним из хорошо исследованных блокаторов Na-каналов является тетродотокснн, который необратимо связывается с белком канала и позволяет его маркировать для последующей очистки. Наибольших успехов в исследовании функции и структурной организации натриевых каналов добились японские исследователи Р. Нума и др. Они показали, что этот мембранный белок представляет собой гликопротеид сМг = 250-300 кД, состоящий из нескольких субъединиц, которые образуют на внутренней поверхности гидрофильную трубчатую структуру, при денатурации в восстановительных условиях белок диссоциирует на два основных компонента, которые специфически связывают Н-тетродотоксин в присутствии фосфолипидов. Диаметр поры этого канала колеблется в пределах 0,4-0,6 нм. Через такую пору могут проходить ионы натрия, связанные с молекулами воды. Избирательность для ионов Na существует, но не является абсолютной.
ТТХ-связывающие белки выделены из различных объектов: головного мозга, клеток нейробластомы, нейронов моллюсков, аксонов кальмара и др. С помощью моно- и поликлональных антител «показано наличие общих антигенных детерминант у белков каналов, выделенных с помощью тетродотоксина. Иммунохимические данные наряду с результатами ограниченного протеолиза и химической модификации молекул свидетельствуют в пользу трансмембранной модели потенциал-независимого натриевого канала. Доступность некоторых участков белка для иммуноглобулинов в липидных мембранах или липосомах подтверждает гипотезу о значительных конформационных перестройках молекулы натриевого канала под действием электрического поля.
В настоящее время установлена полная первичная последовательность ТТХ-чувствительных белков и структура гена, кодирующего синтез в нервной клетке Na-каналов.
Изменение конформационного состояния структурных компонентов ионного канала тесно связано с процессами фосфо-рилирования а- и р-пептидных субъединиц. Обнаружено, что Na-каналы мозга млекопитающих содержат одну а-субъедини-цу, ассоциированную с двумя полипептидами: pj и р2. Установлено, чтоа-субъединица является трансмембранным белком, имеющим участки связывания ряда нейротропных веществ на внешней поверхности мембран, и участки связывания для фосфорилирования цАМФ-зависи-мой протеинкиназой. Оказалось, что этого единственного а-полипептида вполне хватает, чтобы сформировать ионный канал, но не достаточно для выполнения его функций.
Функционированию ионного канала способствуют Рр и р2-субъединицы, которые размещены в основном на внешней поверхности мембран и ковалентно связаны с а-субъединицей через дисульфидную связь. Эти р-субъединицы сильно гликозилиро-ваны, приблизительно 30% их массы составляют карбогидраты, большая часть которых приходится на сиаловую кислоту. Последняя и придает ионному белковому комплексу сильный отрицательный заряд, который и позволяет полноценно функционировать каналу. С другой стороны, эти карбогидраты необходимы ддя нормального биосинтеза и точной сборки функционального канала в нейронах. Показано, что если гликозилирование ингибировано, то новый синтез а-субъединицы быстро останавливается, и она не включается в клеточную поверхность мембраны.
Процесс открывания Na-каналов под влиянием изменения потенциала мембраны — активация натриевых каналов — один из наиболее ярких примеров конформационных перестроек белков под влиянием электрического поля. Открывание каждого канала совершается по известному принципу — "все или ничего". Этот процесс может быть остановлен инактивацией, которая опять-таки связана с переходом белков канала в другое конформационое состояние. Полный цикл активации и инактивации охватывает десятки тысяч натриевых каналов.
3. К-КАНАЛЫ
Потенциал-зависимые калиевые каналы так же, как и натриевые, распространены повсеместно в наружных мембранах нервных клеток и играют столь же важную роль в передаче скоростных сигналов. В отличие от ионов натрия, которые вызывают локальную деполяризацию мембраны и генерирование потенциала действия, калиевые каналы приводят к гиперполяризации нейрона и появлению тормозных потенциалов. Система быстрых калиевых каналов играет большую роль в стабилизации ритмической деятельности нейрона, которая является основным способом кодирования и передачи клеткой химических сигналов. Характерной чертой участия калиевых каналов в ритмической активности является резкое замедление нарастания деполяризации мембраны, вызванной предшествующим входом ионов Na.
Калиевые каналы являются более избирательными для ионов: они не пропускают практически ионы Na, проницаемость для ионов Rb, NH4+ сравнительно мала. Полагают, что селективный фильтр калиевого канала имеет размеры порядка 0,26-0,3 нм. Ионы большего размера не проходят через канал по стерическим причинам, ионы меньшего диаметра — в связи с тем, что они не могут успешно взаимодействовать с кислородо-содержащими анионами, находящимися в боковых цепях гидрофильных аминокислот, которые выстилают внутреннюю белковую пору.
Калиевые каналы подразделяются на три подтипа в зависимости от скорости проведения: быстрые, средние и медленные. Первые два подтипа каналов являются зависимыми от ионов Са и блокируются токсином скорпиона, в то время как третий вид калиевого канала блокируется одним из токсинов ада кобры и пчелы — апамином.
Структура К-каналов в принципе сходна со структурой Na-каналов. На основании данных радиационной инактивации замороженных мембран были определены молекулярные массы для целого олигомерного комплекса — от 165 до 400 кД в зависимости от типа клетки. Обнаружено, что у разных организмов сочетание полипептидных компонентов, составляющих макромолекулу ионофора, существенно различается. В отличие от белков других каналов белки калиевых каналов практически не гликозилированы.
Недавно были проведены работы по выделению генов, кодирующих синтез калиевых канальных полипептидов. Специфическая мРНК, выделенная из мозга крыс, была инъецирована в ооциты лягушки. Показано, что в этом случае регистрируются "новые" калиевые каналы. Найдена высокая степень го-мол огичности между нуклеотидными последовательностями, кодирующими синтез калиевых каналов в разных клетках. Особенно это касалось гидрофобных доменов, которые оказались наиболее консервативными в эволюции.
4. Са-КАНАЛЫ
Транспорт Са+ через кальциевые каналы жизненно важен для разнообразных клеточных функций, особенно в нервной ткани. Электровозбудимые кальциевые каналы изучены преимущественно на нейронах моллюсков. Сейчас становится очевидным, что у высших позвоночных они мало отличаются по физико-химическим характеристикам.
Специфичность кальциевых каналов не очень высока, они способны пропускать из наружной среды Na+ и ионы других щелочных металлов, если концентрация Са+ в наружной среде находится ниже микромолекулярного уровня. Кальциевые каналы пропускают также катионы других двухвалентных металлов, например Mg+ и Мп+. Однако эти катионы легко связываются внешней химической группировкой канала и становятся при определенных концентрациях эффективными блокаторами кальциевого канала. Полагают, что эта группировка является карбоксильной группой, находящейся в устье канала.
Общая схема молекулярной организации кальциевых каналов сходна с описанной выше для Na-каналов. Однако главная а-субъединица окружена большим числом субъединиц, служащих модуляторами активности канала. Пока не ясно, какие химические группировки ответственны за трансмембранный перенос кальция, понятно только, что он - существенно зависит от внутриклеточной концентрации Са+ и функционирования системы циклических нуклеотидов.
Несмотря на то, что численность кальциевых каналов значительно меньше, чем натриевых и калиевых ион-транспортных систем, при определенных условиях они могут самостоятельно вызывать деполяризацию нейрона. Однако сейчас очевидно, что главная функция кальциевых каналов состоит в сопряжении электровозбудимости с внутриклеточными процессами. Эта функция кальциевых каналов особенно важна для включения механизма выхода нейромедиатора из нервного окончания.
5. СИСТЕМЫ АКТИВНОГО ТРАНСПОРТА ИОНОВ. Na+/K+ И Na+/Ca+ - НАСОСЫ
Как уже упоминалось, электрическое возбуждение в нервной ткани существенно зависит от механизмов пассивного и активного мембранного транспорта, контролирующего концентрации ионов и молекул внутри клеток и нередко в межклеточном пространстве. Аксоны обладают большим резервом Кононов и дефицитом Na+-noHOB. Миграции ионов, обеспечивающие прохождение импульсов и создающие изменения потенциала мембраны, быстро компенсируются этими резервами. "Выносливость" аксона очень велика — утомление наступает лишь после прохождения 10 — 10 импульсов, тем не менее перемещения ионов при прохождении импульса должны быть компенсированы в стадии покоя. Кроме того, мембрана в стадии покоя не является абсолютным барьером для перемещений ионов и постепенного уменьшения потенциала.
Ряд внутриклеточных процессов требует постоянной регуляции за счет активных ионных потоков через -мембрану. Наконец, особенно велики нарушения градиентов ионных концентраций при функционировании синапсов. Для компенсации всех этих нарушений градиентов служат ионные насосы.
Основной транспортной системой в нейронах, как и в большинстве других эукариотических клеток, является насос, который вытесняет Na+ и постоянно накапливает К+. Этот процесс требует присутствия АТФ и специфически ингибируется кардиоактивными гликозидами типа оуабаина.
Na+, К+-активируемая аденозинтрифосфотяза, или АТФ-фосфогидролаза К.Ф.3.6Л плазматических мембран нервных клеток осуществляет трансмембранный перенос одновалентных катионов против градиентов их электрохимических потенциалов, используя энергию гидролиза АТФ. Работая с максимальной скоростью, этот ферментативный комплекс способен транспортировать через мембрану около 200 ионов Na и 130 ионов К в 1с. Однако фактическая скорость работы фермента определяется потребностями клетки. У большинства нейронов на поверхностной мембране расположено до 200 натриевых насосов на квадратный микрон, причем в некоторых участках этой поверхности их плотность в 10 раз выше.
Молекула фермента состоит из каталитической а-субъединицы и $-гликопротеида, функциональная роль которого до сих пор неизвестна. Молекулярная масса комплекса белковых субъединиц Na+, К+-АТФазы составляет около 275 000, и размеры фермента колеблются в пределах 6-8 нм. Кроме полипептвдов Na+, К+-АТФаза содержит 3 углеводные цепи, которые присоединены к р-субъединице гликозидными связями. Одновременное использование методов генной инженерии и химии белка привело к установлению первичной структуры Na+, К+-АТФазы. С помощью моноклональных антител установлено внутри- и внеклеточное расположение некоторых участков а- и р-субъединиц. Предложена модель полипептидной цепи фермента, согласно которой сс-субъединица 7 раз пересекает бислойную мембрану и локализована главным образом в цитоплазме. свидетельствует в пользу того, что существуют общие функциональные и структурные взаимодействия между Na+, К+-АТФазой и ионными каналами для транспорта К+.
Сходным с Na+, К+-АТФазой является другой ионный насос — Ка+/Са+-АТФаза. Она производит обмен каждого иона Са+ на 3 иона Na+. Значение этой системы особенно велико в нервных окончаниях, где система медиаторов связана с вхождением Са+ в терминаль и необходимостью компенсировать далее эти смещения градиента. Кроме того, ряд событий в постсинапти-ческой зоне тоже сопряжен с временным вхождением Са+.
6. МОЛЕКУЛЯРНЫЕ ОСНОВЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ, РЕЦЕПТОРЫ
Нейрон способен иметь до нескольких десятков тысяч межклеточных контактов, большинство из которых обеспечивается определенными морфологическими структурами — синапсами. Клеточную поверхность нейронов можно рассматривать как приемник разнообразнейших сигналов.
Чем выше степень эволюционной организации нервной системы, тем разнообразнее природа химических синапсов. Особенно это касается головного мозга высших млекопитающих, включая человека. Очевидно, химические синапсы оказались эволюционно более выгодными для передачи дискретных сигналов по сравнению с другими типами межклеточных контактов, поскольку на их 'основе возможна не только передача сигнала, ко и его разнообразная модуляция, в том числе гуморальными факторами. Основой восприятия нейроном химического сигнала в синапсе, а также ряда модулирующих влияний являются рецепторы.
Рецепторы представляют собой надмолекулярные образования, состоящие из белков, а также гликолипидных компонентов. Они способны под действием медиатора либо нашэсредственно изменять потоки ионов через мембрану, либо индуцировать образование вторичш>тх мессенджеров, которые, в свою очередь, меняют ряд свойста лейрона.
Межнейрональные химические синапсы подразделяются на два типа: возбуждающие и тормозные, причем.первые, как известно, способствуют генерации новых импульсов, а вторые приводят к снятию действия приходящих сигналов. Это деление определяется в значительной мере природой рецепторов. Известны случаи, когда один и тот же медиатор оказывает возбуждающее или тормозное действие в зависимости от природы рецептора.
В зависимости от места положения синапсов их можно подразделить на сомато-аксональные, дентрито-аксональные, ден-трит-дентритные и др. Каждый из этих синапсов имеет свои особенности в функционировании. Схематически структура синапса может быть представлена следующим образом.
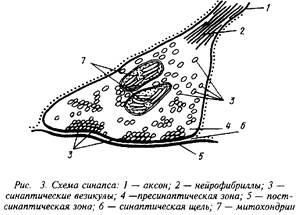
На рисунке хорошо видны утолщения, составляющие пре-синаптическую мембрану подходящего аксона, синаптическая щель и постсинаптическая мембрана. В пресинаптическом окончании находятся синаптические везикулы — хранилища запасов нейромедиатора в пресинаптическом нейроне. Постсинаптическая мембрана является носителем рецепторов. В ряде случаев сами рецепторы могут быть визуализированы при посредстве электронной микроскопии.
При распространении нервного импульса происходит деполяризация пресинаптической мембраны я изменение ионных токов. Наиболее важным событием в нервном окончании является мобилизация ионов Са, которые вызывают миграцию и открывание многочисленных синаптических везикул. Эти везикулы непосредственно связываются с участками пресинапса и открытие их приводит к высвобождению нейромедиатора и диффузии его в синаптическую щель. В терминали аксона сконцентрированы и ферменты синтеза медиатора, митохондрии для энергетического обеспечения этого процесса, системы белков-транспортеров, способствующих узнаванию и обратному захвату молекул нейромедиатора. Этот последний механизм, по-видимому, существенно экономит затраты на синтез готового нейромедиатора и участвует в регуляции срока его действия.
В отличие от биохимических процессов выброса нейроме-диаторов из пресинапса, имеющих общий характер, постсинап-тическое действие нейромедиатора прекращается самыми разнообразными способами: разрушением его определенными ферментами, либо быстрым поглощением из области синапса гли-альными клетками, либо обратным захватом его в пресинаптическую терминаль. В качестве примера существования разных механизмов утилизации нейромедиаторов из синапса можно привести механизмы процессов, происходящих в холинергиче-ском, глутаматергическом и ГАМК-ергическом синапсах.
Механизм разрушения ацетилхолина преимущественно связан с работой фермента — ацетилхолинэстеразы, который располагается на постсинаптической мембране и быстро гидроли-зует медиатор после взаимодействия с рецепторами. В глутаматергическом синапсе механизм удаления нейромедиатора заключается преимущественно в поглощении L-глутамата окружающими глиальными клетками. L-глутамат превращается в глутамин с помощью фермента глутаминазы, находящейся в глиальных клетках. В ГАМК-ергическом синапсе преобладает система обратного захвата медиатора.
7. ОБЩИЕ ВОПРОСЫ НЕЙРОРЕЦЕПТИИ
В биохимии и физиологии нервной системы длительное время доминировала точка зрения, согласно которой местом активных пластических изменений нейронов является пресинапти-ческая мембрана. Были представлены многочисленные свидетельства, касающиеся сдвигов количества квантов нейромедиатора в процессе обучения, памяти, выработки условных рефлексов и др. Постсинаптическим мембранам отводилась либо пассивная роль, либо они вообще не рассматривались в качестве активных участников событий в нервных клетках. Впоследствии стало ясно, что процессы, происходящие в плазматической или синаптической мембране нейрона, являются одними из ключевых для понимания интегративных функций ткани мозга, решения проблем обеспечения эффективного взаимодействия между нервными клетками. В последние десятилетия особое внимание было обращено на изучение структурных компонентов постсинаптических мембран, в частности нейрорецепторов. Исследования тонкой структуры и функции нейрорецепторов показали их важную роль в трансформации химического сигнала в биоэлектрические потенциалы и в передаче информации на внутриклеточные реакции, которые определяют метаболизм нервной ткани.
Следует отметить, что нейрорецепторы расположены как на мембранах нейронов, так и на мембранах глиальных клеток. Однако у последних они имеются в ограниченном наборе и числе. Рецепторные системы, расположенные на глиальных элементах, отличаются от нейрональных весьма важным моментом — они не способны продуцировать ответные реакции клеток в виде оперативных единиц информации. Иными словами, они не генерируют потенциалов действия. Как правило, глиальные клетки реализуют свое действие через внутриклеточные и межклеточные трофические регуляторные реакции, участвуя в метаболизме нейронглия.
Несмотря на огромное разнообразие клеточных рецепторов на мембране нейрона, их можно подразделять на два основных класса, которые различаются по механизмам действия и скорости проведения сигналов. Существуют быстродействующие ионотропные и медленнодействующие метаботропные рецепторы скорость действия первых составляет миллисекунды, в то время как у последних они находятся в секундно-минутном диапазоне. Время действия нейрорецепторов определяется структурной организацией рецепторных компонентов.
Быстродействующие рецепторы содержат в своей структуре ионный канал, открывающийся при контакте с нейромедиатором. Медленнодействующие рецепторы представляют собой комплекс из нескольких белков, которые при воздействии нейромедиатора последовательно меняют конформацию и в конечном счете активируют синтез или выход вторичного, уже внутриклеточного, медиатора. Эти два класса рецепторов обозначают нередко как рецепторы I и II класса. Для правильного восприятия терминологии целесообразно также указать, не рассматривая пока детали, что рецепторы класса II содержат в числе белков, передающих сигнал, так называемые G-белки. Их нередко упоминают, обозначая рецепторы этого класса.

Кроме охарактеризованных выше двух классов рецепторов существуют еще три особые группы рецепторов, которые хотя и присутствуют в нервной системе, но пока представляются не связанными прямо со специфическими функциями последней. К ним относятся рецепторы, переносяшие свои лиганды через мембрану, рецепторы, обладающие собственной тирозинкиназной активностью и, наконец, своеобразная группа, которая при взаимодействии с лигандом претерпевает частичное протеолитическое расщепление. В настоящем руководстве мы не рассматриваем эти группы рецепторов.
К первому классу рецепторов принадлежат никотиновые рецепторы ацетилхолина, рецепторы ГАМКА, глицина, а также часть рецепторов глутамата и аспарагиновой кислоты. Рецепторы катехоламинов, серотонина, ГАМКВ и ряда пептидных соединений, а также мускариновые рецепторы ацетилхолина и некоторые из рецепторов глутамата относят ко второму классу. Последние типы рецепторов через систему вторичных посредников вызывают изменения в активности проте-инкиназ, способных фосфорилировать мембранные белки, включая ионные каналы.
Таблица 1
Структура, общий характер и функции рецепторов класса I
| Тип рецептора | Преимуществен -ная функция | Мг, кД | Субъединицы |
| Холинергический | Возбуждающая | 250 | а 40-50 Р 50-54 у 56-60 6 58-65 |
| ГАМКА | Тормозная | 230-260 | |
| Глутаматный | Возбуждающая | 240-400 | 90-10 |
| Глициновый | Тормозная | 246 | а 48 Р 58 |
Таблица 2
Масса белков рецепторов класса II
| Рецептор | Мг, кД |
| Ад рене р ги чески й | 58-80 |
| Глутаматер гически й | 90-110 |
| Холинергический | 85-105 |
| Дофаминовый | 72-94 |
| Опиатный | 53-65 |
| Серотониновьш | 67 |
Следует отметить, что в последние годы обнаружена группа нейрорецепторов, связь которых с ионными каналами осуществляется через G-белки, не сопряженные с перечисленными выше вторичными мессенджерами. Хотя в такую систему рецепции и не включены протеинкиназы, тем не менее участие G-белка в трансформации сигнала значительно увеличивает время действия по сравнению с нейрорецепторами класса 1.
Фундаментальным свойством всех нейрорецепторов является их лабильность и высокая скорость синтеза самого рецептора. Это свойство рецепторов конрастирует с более жесткой запрограммированностью синтеза белковых компонентов мембран, которая обычно наблюдается у других типов тканей. В нейронах развиты механизмы непрерывного синтеза рецепторов и их быстрой утилизации либо путем интернализации, либо с помощью пиноцитоза. Высокая скорость обновления нейрорецепторов обусловлена, по-видимому, необходимостью изменения "информационной емкости и "пропускной способности" нейрона. В этом случае генетический аппарат клетки способен, интегрируя всю приходящую информацию, "принять решение" путем перестройки синтеза белковых компонентов мембран. В этом скрыта одна из причин уникального свойства нейронов и нервной ткани в целом — пластичности.
Таким образом, основная роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов.
Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют альтернативу. С одной стороны, это изменение чувствительности к рецептору, с другой — увеличение или снижение количества активных рецепторов на мембране. Заслуживают внимания и гипотезы, касающиеся посттрансляционной модификации нейрорецепторов, которая позволяет изменить количественные параметры их функционирования.
Внимание к проблемам нейрорецептии со стороны биохимиков, фармакологов и физиологов обусловлено еще и тем, что причиной многих дисфункций нервной системы является нарушение целостности мембранных компонентов как нейронов, так и глиальных клеток. Отметим, что существующие успехи в лечении некоторых нервно-психических заболеваний связаны в большей мере с прогрессом в исследовании именно молекулярных свойств ряда рецепторов. Оказалось, что многие нейрорецепторы выполняют роль избирательных мишеней действия известных лекарственных препаратов. Исследования в этой области нейробиологии служат сейчас постоянным источником для целенаправленного поиска и создания новых классов фармакологических средств, обладающих улучшенными терапевтическими свойствами.
8. БИОХИМИЧЕСКИЕ МЕТОДЫ ИЗУЧЕНИЯ МЕХАНИЗМОВ НЕЙРОРЕЦЕПТИИ
Наиболее широко распространенным и разработанным методическим подходом для количественного анализа взаимодействия нейромедиаторов со своими рецепторами на мембране клетки является радиолигандный метод. Суть этого метода заключается в изучении параметров связывания радиоактивного лиганда с мембранно-связанными или изолированными рецепторными белками. В настоящее время существует хорошо развитая кинетическая теория рецептии и методы определения физико-химических параметров процесса образования комплекса лиганд-рецептор. Такой физико-химический анализ позволяет сделать определенные заключения о структуре активных центров нейрорецепторов, в частности, выяснить природу некоторых функциональных групп, которые ответственны за первую стадию взаимодействия лиганда с акцептором.
Для того чтобы кратко ознакомиться с количественной теорией взаимодействия веществ со своими рецепторами, рассмотрим простейшие условия, когда одна молекула лиганда взаимодействует с одним центром связывания:

где L — лиганд; Q — центр связывания; В — комплекс лиганда со связывающим центром; К{ и K.j — кинетические константы. При динамическом равновесии скорость реакции образования комплекса В равна его скорости диссоциации, т.е. Vt = V_1? тогда концентрация вычисляется по формуле
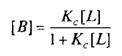
При этом предполагается, что L и Q взаимодействуют между собой по закону действующих масс, т.е. скорости реакций образования комплекса и его диссоциации прямо пропорциональны концентрациям компонентов в системе. Отношение констант прямой и обратной реакции называют константой сродства Кс. Она характеризует соотношение занятых и свободных участков связывания при данной концентрации лиганда. Обычно для описания параметров связывания используют величину, обратную константе сродства,—
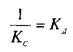
константу диссоциации. Эта константа соответствует величине, при которой происходит насыщение 50% связывающих участков:

Если вместо константы сродства Кс использовать обратную ей величину Кд, то, подставив это значение в уравнение, характеризующее равновесную реакцию взаимодействия лиганда с рецептором, получим следующее уравнение:

Приняв общее число рецепторов за 1, можно преобразовать уравнение к виду, аналогичному уравнению Михаэлиса, которое используется в энзимологии для описания кинетики обратимых ферментативных реакций:

где концентрация комплекса фермент-субстрат; —
концентрация субстрата и Ks константа диссоциации комплекса; — исходная концентрация субстрата.
Согласно этим уравнениям зависимость величины эффекта от дозы лиганда или фермента описывается гиперболой. Чаще всего для работы пользуются графическим выражением зависимости эффекта не от концентрации, а от логарифма концентрации лиганда. Графически зависимость результатов может быть представлена разными способами, однако наиболее информативным способом расчета являются координаты Скэтчарда. Действительно, помимо равновесной константы связывания и общей концентрации центров связывания этот метод позволяет определить концентрацию свободного лиганда, соответствующую данной концентрации комплекса В. Константа диссоциации равна котангенсу угла наклона прямой. Отрезок на оси абсцисс от точки пересечения с прямой до начала координат соответствует максимальному уровню насыщения центров связывания.

Таким образом, представление результатов равновесного связывания в координатах Скэтчарда дает информацию о характере протекающего процесса и позволяет определить важные параметры лиганд-рецепторного взаимодействия — константу диссоциации и концентрацию центров, способных образовывать комплексы с нейромедиатором.
В качестве примера изучения рецепторного связывания нейромедиатора с белковыми компонентами на мембране нейрона приведем экспериментальные исследования глутаматных рецепторов радиолигандным методом. Так, исследования параметров связывания Н-глутамата с синаптическими мембранами, выделенными из коры больших полушарий головного мозга крыс, показали их зависимость от чистоты материала, способов хранения, условий проведения реакции связывания и др. При стандартизации всех указанных условий зависимость специфического связывания Н-глутамата с синаптическими мембранами имеет насыщающий характер. Представление экспериментальных данных в координатах Скэтчарда свидетельствует о наличии на мембранах однородной популяции участков связывания с Кд - 89,4 нМ и Вмакс = 2,0 пмоль/мг белка.

Значение количества центров связывания, выраженное в СРМ, пересчитывается в фмоль/мг белка по следующей формуле:
 >
>
где А^ — молярная активность радиолиганда, Кю/моль; а — 2,210~ pacn/мин; f — эффективность счета; — разность счета связывания радиолиганда с рецептором в отсутствие и в присутствии немеченого радиолиганда; t время счета; С — концентрация белка, мг.
Для того чтобы отличить эти параметры связывания от неспецифического связывания и поглощения глутамата другими участками мембраны, существуют дополнительные экспериментальные приемы, в том числе проведение реакции в присутствии разных катионов. Истинное рецепторное связывание глутамата является Na*-независимым процессом, в то время как поглощение и транспорт этого нейромедиатора другими участками синапса происходит в присутствии высоких концентраций ионов Na.
Далее возникает вопрос, соответствуют ли эти независимые участки связывания самого глутамата тем рецепторным компонентам на мембране нейрона, которые способны вызывать физиологический ответ клетки на данный медиатор. Оказалось, что сродство и константа диссоциации, полученные экспериментальным биохимическим методом, находятся в пределах физиологических концентраций действия L-глутамата на нейроны позвоночных. Такие показатели реакции связывания нейромедиатора, как насыщаемость и обратимость, соответствуют аналогичным свойствам глутаматного рецептора, регистрируемым с помощью электрофизиологических методов. Более того, чувствительность к ряду известных агонистов и антагонистов, таких как NMDA, каинат, квисквалат и другие, была сходна с физиологическими ответами. Следует упомянуть, что характер связывания нейромедиатора в присутствии ионов Na существенно отличается от рецепторного взаимодействия и коррелирует с параметрами высокоаффинного поглощения L-глутамата клетками, регистрируемыми физиологически. Все это иллюстрирует пути оценки параметров связывания нейромедиатора и специфические трудности, возникающие при такой оценке.
Одним из основных подходов к изучению молекулярных свойств нейрорецепторов является изолирование индивидуальных рецепторных белков, специфически связывающих нейромедиаторы или необратимо взаимодействующих с их антагонистами или бло-каторами. Так, прогресс в исследовании никотиновых холино-рецепторов был обусловлен обнаружением а-бунгаротоксииа, который оказался специфическим блокатором этого типа рецепторов и позволил выделить мембранные белки и очистить их на основе радиолигандного метода. Наличие таких приемов дает возможность разграничить хеморецепторные процессы от ферментативного и транспортного метаболизма нейромедиато-ров. Особенно это важно для изучения рецепторов аминокислотных медиаторов нервной ткани.
Изучение химической природы мембранных белков включает предварительное выделение, солюбилизацию, очистку и анализ очишенных компонентов. Причем применение классических методов структурного анализа для характеристики мембранных белков имеет свои сложности и особенности. Как правило, они обусловлены свойствами мембран и их компонентов, в частности, наличием липидных и гликолипид-ных структур. Проблемы, связанные с экстракцией белковых компонентов мембран, их очисткой и анализом, составляют специальный раздел мембранологии. Здесь будут рассмотрены лишь самые общие моменты.
Выбор метода солюбилизации зависит от цели исследования и имеет смысл только тогда, когда дает возможность сохранить нативные свойства рецепторного белка и исследовать его с помощью обычных биохимических подходов. Поэтому выбор со-любилизирующего агента на первом этапе может оказаться ключевым для анализа структуры и функции рецептора.
Существует целый ряд самых разнообразных солюбилизи-рующих агентов, пригодных для решения проблем мембранной биохимии. Наиболее надежными среди них являются неионные и ионные детергенты. В основе их действия лежит амфифиль-ная природа этих агентов, позволяющая им взаимодействовать и с гидрофильными, и с гидрофобными участками мембранных белков. Эффект детергента, разрушающего взаимосвязи в мембране, определяется двумя видами взаимодействия: детергент-белок и детергент—детергент. Большое значение имеет последнее взаимодействие, так как чем выше способность молекул детергента взаимодействовать друг с другом, тем меньше будет количество молекул, способных взаимодействовать с белками. Этот критерий мицеллообразования служит характеристикой детергента и его способности растворять те или иные белковые компоненты. Низкий коэффициент мицеллообразования характерен для мягких солюбилизирующих агентов, таких как тритон Х=Ю0, дезоксихолат натрия, дигитонин и другие, которые позволяют выделять нативные мембранные белки с сохранением их биологической активности. В то же время додецилсульфат натрия с высоким коэффициентом мицеллообразования обладает большой связывающей способностью и значительно повреждает нативную конформацию белков. Как правило, этот детергент используется при анализе субъединичной структуры макромолекул, так как легко разрушает межмолекулярные связи. Это свойство нередко применяется для определения молекулярной массы субъединиц белков при электрофорезе в присутствии ДСН.
Перед тем как приступить к дальнейшему выделению и изучению мембранных рецепторных белков, следует по возможности более полно удалить избыток детергента, поскольку он может оказывать нежелательное действие на биологическую активность и последующий физико-химический анализ структуры нейрорецептора.
Классические методы исследования мембранных белков, в том числе нейрорецепторов, включают практически все биохимические методы с учетом присутствия детергентов. Основным приемом специфического выделения ничтожно малых количеств нейрорецепторов является аффинная хроматография, которая позволила добиться впечатляющих успехов в изучении молекулярных свойств самых разнообразных типов нейрорецепторов.
Эффективность аффинной хроматографии зависит преимущественно от выбора лиганда или акцептора, который определяет природу выделяемого мембранного белка. Существенным фактором в этом случае является сродство лиганда к рецептору, и поэтому самыми эффективными лигандами оказываются специфические блокаторы или антагонисты нейрорецепторных белков. Иногда для выделения конкретного белка используют две или три ступени аффинной хроматографии на разных сорбентах и с разными лигандами. Получили широкое распространение методы иммуноаффинной хроматографии, в которых в качестве лиганда используется поликлональные или моноклональные антитела, полученные к компонентам рецептора.
Дальнейшее выделение и разделение фракций обычно осуществляют с помощью высокоэффективной жидкостной хроматографии, которая позволяет очищать индивидуальные компоненты мембранных белков. Причем обратнофазная хроматография дает уникальные возможности по разделению гидрофобных белков и пептидов. Нативность белковых компонентов рецепторов проверяют либо по лигандсвязьшающей функции, либо путем реконструкции их функции в разных модельных системах.
Одной из таких модельных систем, позволяющих контролировать ионтранспортные или ионселективные функции нейрорецепторов, служат липосомы. Способность липосом встраивать белки или целые рецепторные комплексы с сохранением их функциональной активности используется в мембранологии для моделирования функций белков "в чистом виде". В этом случае можно получать информацию о структурной организации компонентов, составляющих макромолекулу рецептора, и их внутренних перестройках в контролируемых условиях эксперимента.
В настоящее время разработано большое количество методов получения липосом, которые могут изменять фосфолипидный состав, заряд, "текучесть" или многослойность их компонентов. Размеры липосом могут варьировать от 25 нм до 100 мкм. Функцию белков, встраиваемых в липосомы, контролируют по динамике транспорта или накопления меченых ионов внутри липосом.
В последние годы исследователи возлагают особые надежды на иммунохимические способы идентификации структурных компонентов нейрорецепторов. Высокая специфичность антител и их способность узнавать разные антигенные детерминанты рецепторных комплексов широко используется для выяснения структурной организации нейрорецепторов и процессов их биосинтеза, включая генно-инженерные исследования. Иными словами, поли- и моноклональные антитела являются важным инструментом для изучения механизмов рецептии и общих вопросов нейробиологии.
Принципиальным решением множества проблем, связанных с применением антител, явилось создание новой гибридомной технологии, которая позволила получить моноклональные антитела. Эта техника была разработана в 1975 г. У.Келлером и А.Милстейном. Получаемые с помощью этого метода гибридные клетки синтезируют и выделяют в культуральную среду антитела, абсолютно одинаковые по своему сродству к той или иной антигенной детерминанте.
Гибридомная техника позволила получать самые разнообразные моноклональные антитела против химически индивидуальных антигенных детерминант на одной молекуле белка. В настоящее время моноклональные антитела широко используют для идентификации практически любых макромолекул, включая нейрорецепторы.
Следует подчеркнуть, что изучение структуры и функции нейрорецепторов и других мембранных компонентов нейрона является не самоцелью для нейробиологической науки. Важно понять, как молекулярные процессы, происходящие в нервных клетках, способны интегрировать самую разнообразную информацию и реализовать ее в виде сложных поведенческих, высших психических и эмоциональных реакций. Этот стратегический путь "от простого к сложному" получил в последние годы мощный импульс благодаря разработке принципиально новых способов прижизненной регистрации динамических биохимических реакций, происходящих в клетках головного мозга. Появились методы позитронно-эмиссионной томографии, ядерно-магнитного резонанса, гамма-сцинциграфии и другие, позволяющие прижизненно регистрировать системный метаболизм разных органов и тканей, включая головной мозг млекопитающих. Это создает предпосылки для успешного изучения нейрохимических основ формирования разнообразных функциональных состояний живого мозга человека, его наиболее сложной сферы деятельности —психической.
Одним из информативных методов является позитронно-эмиссионная томография, суть которой сводится к регистрации специальным устройством радионуклидных маркеров меченых химических соединений, включающихся специфически в тот или иной метаболический процесс. Причем этот процесс может быть воспроизведен в виде томографических, т.е. объемных послойных изображений распределения позитронной метки по структурам и зонам головного мозга. Наиболее точная локализация достигается при использовании одновременно двух противостоящих детекторов, регистрирующих совпадающие лучи. В настоящее время уже существует хорошо разработанные приемы оценки функционального состояния головного мозга с помощью измерения локального метаболизма глюкозы, медиаторов, С02 и кислорода в процессе разнообразной деятельности индивида.
Важной особенностью метода, позволяющей его использовать в прижизненном исследовании деятельности мозга людей, является применение изотопов, излучения которых с учетом сроков распада безвредны для организма.
Сейчас получены четко различающиеся "карты" излучений при различных формах деятельности мозга человека, например восприятия слов, обдумывания слов, воспроизведения энграмм и др. Резкие различия регистрируются при воздействиях на мозг наркотиков и других психотропных агентов.
Естественно, что при дальнейшей разработке метода ПЭТ и его внедрении в клиническую нейробиологию возник вопрос о выборе адекватных маркеров, которые способны выявлять си-наптические реакции в нервной ткани. Ряд исследователей успешно работают с препаратами, которые позволяют визуализировать определенные нейрорецепторы и выявлять конкретные медиаторные пути, включающиеся в выполнение того или иного вида деятельности мозга человека. В клинике этот метод дает возможность проводить раннюю диагностику не только опухолевых новообразований, но и контролировать различные деструктивные процессы. Кроме того, применяя его, можно определять эффективность лечебного воздействия фармакологических средств и их правильный выбор для успешного лечения болезней мозга. В качестве иллюстрации к сказанному следует привести исследования, проводимые Вагнером и его коллегами по изучению вклада дофаминергических путей и их рецепторов в патогенез некоторых заболеваний. Выбор дофаминовых рецепторов был обусловлен их четкой локализацией в некоторых подкорковых экстрапирамидных структурах и известной их дисфункцией при двигательных расстройствах, паркинсонизме и шизофрении.
Предпосылкой для применения в ПЭТ агонистов и антагонистов дофаминовых рецепторов явились результаты радиоли-гандного связывания известных аналогов дофамина с синаптическими мембранами in vitro. Параметры связывания, константы диссоциации и количество связывающих участков сопоставляли с данными, полученными при ПЭТ, так как связывание радиофармпрепаратов с мембранами клеток, головного мозга in vivo имеет аналогичные закономерности. Расчет прижизненного взаимодействия нейрорецептор-лиганд имеет некоторые особенности, однако они учитываются непосредственно в программах компьютерного обеспечения. Наиболее удачными радио-лигандами для исследования дофаминовых рецепторов оказались антагонисты пС-метилспиперон и лУС-спироперидол. Поглощение и селективная избирательность накопления этих соединений в базальных ганглиях головного мозга коррелировали со степенью деструктивного процесса у больных паркинсонизмом. Эти исследования подтвердили гипотезу о первичной вовлеченности нигро-стриарных структур в регуляцию двигательных функций.
Дальнейшее развитие регистрирующих устройств ПЭТ, увеличение степени разрешения приборов на микроуровне и поиск новых избирательных радиофармпрепаратов могут открыть уникальные перспективы прижизненного изучения динамических биохимических процессов, происходящих в ткани мозга при выполнении сложных видов деятельности.
9. ОБЩИЕ ХАРАКТЕРИСТИКИ СТРУКТУРЫ И ФУНКЦИИ ИОНОТРОПНЫХ РЕЦЕПТОРОВ
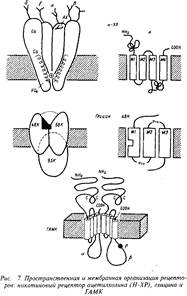
Основой всех ионотропных рецепторов является крупный белок, состоящий из пяти, реже четырех, белковых субъединиц. Молекулярные массы субъединиц варьируют обычно в пределах от 40 до 70 кД. Первичная структура белков различных ионотропных рецепторов обнаруживает высокую степень гомологии — от 20 до 60%, что указывает на общность эволюционного происхождения. Субъединицы рецептора пронизывают толщу клеточной мембраны, образуя ионный канал. Участки полипептидных полей субъединиц, выстоявшие над поверхностью клетки, служат для узнавания и взаимодействия с медиатором. Участки субъединиц, проходящие через толщу фосфолипидной мембраны и образующие собственно канал, характеризуются богатством гидрофобных неполярных аминокислотных остатков, обладающих высоким сродством к липидному окружению рецептора. Участки субъединиц, расположенные на внутренней поверхности мембраны, служат, во-первых, для взаимодействия с клеточными скелетными белками, ограничивающими их подвижность, и, во-вторых, являются мишенью для факторов, регулирующих активность рецептора в зависимости от ряда внутриклеточных процессов. Лучшим примером ионотропного рецептора служит рецептор ацетилхолина, представленный на рис. 7. Выстоящие над мембраной участки ионотропных рецепторов связаны нередко с углеводными компонентами.
Ионотропные рецепторы, например рецепторы гамма-ами-номасляной кислоты типа А, способны образовать большое количество подтипов за счет различного сочетания субъединиц. Существует более двух десятков подтипов ГАМКд-рецепторов благодаря различным комбинациям а-, р-, у- и5~субъединиц. В состоянии покоя каналы ионотропных рецепторов закрыты. При взаимодействии с медиатором происходит конформапионная перестройка субъединиц рецепторов и каналы открываются на несколько миллисекунду.
После активации рецепторные макромолекулы теряют на некоторое время чувствительность к медиатору. Наступает временная десенситизация.
Природа ионов, которые способен пропускать рецептор, определяется диаметром канала и характеристиками боковых радикалов аминокислотных остатков стенки канала. Никотиновые рецепторы ацетилхолина открывают дорогу ионам К* из клетки и ионам Na+ внутрь клетки; NMDA-глутаматные рецепторы наряду с одновалентными ионами, открывают путь внутрь клетки ионам Са+; ГАМКА и глициновые рецепторы пропускают внутрь клетки С1-ионы.
10. ОБЩАЯ ХАРАКТЕРИСТИКА МЕТАБОТРОПНЫХ МЕДЛЕННЫХ РЕЦЕПТОРОВ
Метаботропные рецепторы представляют собой сложную систему, состоящую, по крайней мере, из трех белков: 1) собственно рецепторного белка, связывающегося с нейромедиатором; 2) так называемого G-белка, модифицирующего и передающего сигнал с рецепторного белка и 3) белка-эффектора, который является ферментом, катализирующим образование внутриклеточного низкомолекулярного регулятора, так называемого вторичного мессенджера. Пример метаботропного рецептора представлен на рис. 8.
Собственно рецепторный белок — R-белок — представляет собой крупный полипептид, состоящий из 400-2 ООО аминокислотных остатков, N-конец пептида выстоит над поверхностью клетки, С-конец направлен внутрь клетки. Пептидная цепь семь раз пересекает клеточную мембрану, образуя соответственно по три петли над и под поверхностью мембраны. N-концевая последовательность нередко гликозилирована. Те части пептида, которые пронизывают фосфолипидную мембрану, состоят в значительной мере из неполярных аминокислот и как бы плавают в липидном слое. Медиатор, вступая во взаимодействие с внешними участками полипептида, меняет их конформацию и, в свою очередь, меняет положение плавающих внутри мембраны участков пептида. В конечном счете это ведет и к изменению конформации участков пептида, находящихся под мембраной. В этой конформации они приобретают способность контактировать со следующим белком комплекса — G-белком. Заметим, что активированный медиатором R-белок способен контактировать последовательно с многими десятками и сотнями молекул G-белка, переведя их, в свою очередь, в активное состояние. Иначе говоря, уже на этой стадии происходит усиление, амплификация сигнала.

G-белок представляет собой олигомер, состоящий из 2-3 субъединиц с общей молекулярной массой порядка 60-100 кД, В неактивном состоянии G-белок обычно связан с молекулой ГДФ. При взаимодействии с активированным R-белком конфигурация G-белка меняется таким образом, что на место ГДФ становится ГТФ. Именно в состоянии комплекса с ГТФ G-белок способен быть активатором следующего компонента системы — фермента, образующего вторичный мессенджер. Активное состояние белка G-белка ограничено во времени тем, что связанные с ним ГТФ расщепляются до ГДФ, и G-белок при этом возвращается в исходное неактивное состояние. Расщепление ГТФ до ГДФ осуществляется самим G-белком, который является, как бы по совместительству, гуанозинтрифосфотазой.
Будучи в активном состоянии, G-белок активирует фермент. Этим ферментом может быть аденилатциклаза, катализирующая синтез цАМФ из АТФ, гуанилатциклаза, катализирующая синтез цГМФ из ГТФ, фосфолипаза С, отщепляющая фосфои-нозитол от фосфоинозитида мембраны. Активированный G-белок может выступать не только в роли фактора, усиливающего действие циклаз, но и в качестве их специфического ингибитора. Разновидности G-белка, выполняющие одну из этих функций, имеют обозначения: Gg-белок, стимулирующий активность циклаз, Gj-белок, подавляющий активность циклаз, и, наконец, О0-белок, активирующий фосфолипазу С. Это лишь главные и наиболее изученные разновидности G-белков. Вообще их число значительно больше. Отметим особо разновидности G-белков, которые, получив сигнал от метаботропного рецептора, передают его на тот или иной ионный канал. Строго говоря, это уже не метаботропный путь, а особая форма включения ионного канала. В отличие от ионотропных рецепторов здесь может, по-видимому, быть достигнута большая продолжительность действия и охват большего числа ионных каналов.
Так же как и стадии передачи сигнала с R-белка на G-белок, стадия активации фермента и стадия синтеза ферментом вторичных мессенджеров сопровождается дальнейшим усилением сигнала.
8. ХАРАКТЕРИСТИКИ ОТДЕЛЬНЫХ РЕЦЕПТОРНЫХ СИСТЕМ
Большинство известных в настоящее время нейромедиаторов участвуют в передаче сигнала и через ионотропные, и через метаботропные рецепторы. Это заставляет строить дальнейшее изложение, следуя перечню медиаторов.
Ацетилхолиновые рецепторы. Ацетилхолин как нейромедиатор периферической и центральной нервной системы взаимодействует с двумя видами холинорецепторов: мускариновыми и никотиновыми. Эти подтипы рецепторов отличаются по специфичности взаимодействия с рядом агонистов и антагонистов ацетилхолина. Так, м-ХР избирательно возбуждаются мускарином, а н-ХР.отвечают на аппликацию никотина. Физиологически важным различием между м-ХР и н-ХР является скорость ответа на приходящий сигнал. Считают, что н-ХР предназначены опосредствовать быстрые и непродолжительные эффекты, в то время как м-ХР реагирует более медленно и длительно.
Никотиновые холинорецепторы. н-ХР оказались более изученными биохимически благодаря существованию двух факторов: наличие специфического нейротоксина, способного блокировать функцию рецептора, и обнаружению большого количества этого рецептора в электрических органах рыб. Структура представлена на рис. 7.
Н-ХР содержит 5 субъединиц: две а-субьединицы с Мг = 40 кД, одну р-субъединицу — Мг = 49 кД, одну у-субъединицу — 60 кД и одну 5-субъединицу — Мг = 67 кД. Катионные группы двух молекул ацетилхолина связываются с анионными участками ct-субъединиц. KD взаимодействия АХ с рецептором близко к 10~ М. Открывающийся при контакте с АХ на несколько миллисекунд канал успевает пропустить до 5-Ю ионов К+ и Na+. АХ, диссоциировавший с рецептором, или "избыточный" АХ в синаптической щели быстро расщепляется ферментом ацетилхолинэстеразой, расположенной на постсинаптической мембране в непосредственной близости от рецептора. Ацетилхолинэстераза является одним из самых быстродействующих, высокооборотных ферментов. Таким образом, сигнал резко ограничен во времени. Образовавшийся холин захватывается белками-транспортерами пресинаптической мембраны и служит далее для ресинтеза АХ в терминал и.
Активность рецептора может модулироваться со стороны клетки фосфорилированием отдельных аминокислотных остатков участка, обращенного внутрь клетки. Подвижность рецептора ограничена связью с цитоскелетными белками через так называемый белок 43К.
По характеру влияния веществ на функцию ХР можно выделить: агонистпы, антагонисты и блокаторы. Наиболее известные из них представлены в табл. 3.
Таблица 8Л
Лиганды холинорецепторов
| Типы лигандов | ||
| никотиновые | мускариновые | |
|
Агонисты Антагонисты, Блокаторы |
Ацетилхолин, карбахол, никотин, лобелии, 1,1 -диметил-4-фенил-пиперазин Гексаметоний, декамстоний а - Бунгаротоксин 6 - Тубокурарин |
Ацетилхолин, карбахол, метахолин, мускарин, диметил ацетилхолин Атропин, галл амин, платифиллин |
Способность разных соединений взаимодействовать с этими рецепторами имеет не только теоретическое, но и большое практическое значение. Поскольку нарушение холинергической медиации лежит в основе ряда патогенетических механизмов заболеваний нервной, эндокринной, иммунной систем, то поиск лекарственных веществ, непосредственно воздействующих на пострецепторные механизмы, является эффективным. Действительно, как показала практика, многие используемые в клинике н-холинергические фармпрепараты имеют точкой своего приложения периферические моторные синапсы, ганглии и некоторые хемочувствительные структуры висцеральных систем. Часто в клинике применяют блокаторы н-ХР, среди которых выделяют ганглиоблокаторы и миорелаксанты. Нарушение функции никотиновых ХР лежит в основе тяжелого прогрессирующего заболевания — миастении гравис. Болезнь резко снижает эффективность нервно-мышечных соединений и обусловлена появлением аутоантител к ХР. Показательно, что содержание аутоантител к ХР в крови больных коррелирует с клиническим состоянием: более высокие титры аутоантител наблюдаются у тяжелых больных миастенией. Лечение этой категории больных с помощью иммунодепрессантов вызывает длительные положительные сдвиги в клинической картине заболевания.
Мускариновые рецепторы ацетилхолина. Эта категория рецепторов ацетилхолина относится к категории метаботропных. Общие их характеристики уже описаны выше. Структурная модель м-ХР представлена на рис. 8. Пептидная цепь, образующая основу м-ХР, состоит из 800-950 аминокислотных остатков и связана с углеводными компонентами. Существует большое число подтипов м-ХР, связанных с различными физиологическими эффектами. В частности, рецепторы м2-ХР, локализованные преимущественно в ЦНС и сердце, продуцируют в качестве вторичных мессенджеров цГМФ и подавляют аденилатциклазу. Рецепторы подтипа МрХР, расположенные, в частности, в желудке и симпатических ганглиях, индуцируют образование инозитолфосфатов и диацилглицерола, которые, в свою очередь, ведут к повышению в цитозоле концентрации Са+ и активации протеинкиназы С. Вторичное модулирующее воздействие они оказывают на кальциевые каналы. Различия рецепторов м1 и м2 выражается также в высоком и низком, соответственно, сродстве к ацетилхолину. Последовательность многообразных молекулярных событий, ведущих к ответу клетки на действие лигандов для м-ХР, показана на схеме 1.

Мускариновые рецепторы АХ связаны с психоэмоциональным восприятием, секрецией слюнных и желудочных желез, функцией сердечно-сосудистой системы и др. Антагонисты м-ХР применяются в клинике в качестве язвазаживляющих, спазмолитических препаратов, а также используются для симптоматического лечения паркинсонизма. В качестве успокаивающих средств, транквилизаторов находят применение в терапии амизил и метамизыл центральные блсекаторы м-ХР.
Таблица 4.
Специфические агонисты и антагонисты ШМК-рецепторов
| Типы лигандов | Типы ГАМК-рецепторов | |
| ГАМКА | ГАМКВ | |
| Агонисты | Гамма-аминомасляная кислота, мусцимол | .Гамма-аминомасляная кислота, баклофен |
| Антагонисты | Бикукулин, пикротоксин | Дельта-амкно валериановая кислота |
Наконец, мощное влияние на активность холи-нергической передачи оказывают агенты, ингибирующие холи-нестеразу и повышающие тем самым концентрацию АХ в си-наптической щели. Таков механизм действия эзерина и его аналогов, применяемых для снижения внутриглазного давления.
ГАМК-рецепторы. Успехи в идентификации различных типов ГАМК-рецепторов, их биологических и фармакологических характеристик тесно связаны с созданием специфических аго-нистов и антагонистов. По локализации ГАМК-рецепторы подразделяются на центральные и периферические, пре-и постсинаптические. Различают два типа рецепторов ГАМК: бикукулин-чувствительные и баклофен-чувствительные. Наиболее изученным является первый тип рецепторов, который чувствителен также к антагонисту пикротоксинину. Обнаружено, что этот тип рецепторов является быстродействующим и сопряжен с ионными каналами для СР. Другой тип ГАМК-рецепторов относится к медленнодействующим рецепторам, и полагают, что он через G-белок ассоциирован с каналами для ионов К+ и Са+.
Исследования физико-химических свойств очищенного ГАМКд-белка показали, что Мг его находится в пределах 220-270 кди что он представляет собой пентамер гликопротеидли-пидной природы, образующий каналы для ионов хлора.
Особенностью ГАМКА-рецепторов является то, что они содержат специфические участки связывания не только самой ГАМК, но и других физиологически активных соединений. Наиболее интересными и изученными среди них являются лекарственные соединения, объединенные под названием бензо-диазепины а также эндогенные регуляторы пептидной природы эндозепины.
Среди лекарственных веществ бензодиазепины занимают особое место в связи с их широким лечебным спектром действия: противосудорожного, снотворного, нейротропного, анти-ксиолитического и др.
Важной особенностью функционирования ГАМК-ергической трансмиссии является система удаления выполнившего свою функцию или избыточного лиганда из синаптической щели. В отличие от многих других синаптических систем ГАМК преимущественно претерпевает обратный захват и возвращается в нервные окончания с помощью белков-транспортеров. Они расположены на пресинаптической мембране и несколько похожи по особенностям структуры на метаботропные рецепторы: пептидная цепь, многократно пересекающая мембрану с образованием системы петель над и под мембраной. Белок-транспортер узнает, захватывает и за счет энергии протонного насоса '.переносит ГАМК внутрь терминали.
Места специфического связывания бензодиазепинов находятся на молекулах белка, входящего в структуру рецептора ГАМК. Активация ГАМКА-рецепторов приводит к открытию ионного канала для хлора, а бензодиазепины.при этом удлиняют продолжительность существования открытых ионных каналов, не влияя на их число и скорость транспорта хлора.
Установлено, что участок связывания бензодиазепинов взаимодействует также с эндогенными пептидными регуляторами — эндозепинами. Последние обладают физиологическими эффектами, противоположными бензодиазепинам, — вызывают возбуждение, тревожность и проконфликтное поведение животных. Они подавляют открытие канала для CP, индуцируемое ГАМК, т.е. являются ее эндогенными функциональными антагонистами. Таким образом, бензодиазепины оказались блокаторами участка связывания эвдозепинов, т.е., так сказать, экзогенными антагонистами эндогенных антагонистов ГАМК. Понятно поэтому, что эндозепины обозначают иногда аббревиатурой DBI.
Имеются указания на существование в ЦНС еще одной категории эндогенных антагонистов ГАМК — производных р-кар-болинов. Они также вызывают тревожность, панические состояния у животных и человека.
Что касается второго типа ГАМК-рецетгоров — ГАМКВ, то кроме отмеченных выше особенностей агонистов и антагонистов они характеризуются преимущественно пресинаптической локализацией и сопряженностью с калиевыми, а не с хлор-каналами; локализованы они главным образом.в периферической нервной системе.
Глициновые рецепторы. Радиолигандные исследования позволили локализовать и изучить особенности распределения в центральной нервной системе участков связывания, которые метятся Н-стрихнином. Эти участки, имеющие Кд = 10~ М, являются рецепторами глицина. Наибольшая плотность глициновых рецепторов обнаружена в области ядер.подъязычного и тройничного нервов, локализованных в продолговатом мозге.
Участки связывания стрихнина найдены также в ретикулярных ядрах продолговатого мозга, моста и среднего мозга. Серое вещество спинного мозга также отличается высокой плотностью глициновых рецепторов как в передних, так и в задних рогах.
Глициновый рецептор спинного мозга млекопитающих был очищен с помощью аффинной хроматографии на аминострих-нин-агарозе. Обнаружено, что он представляет собой гликопро-теид-липидный комплекс с Мг = 250 кД, состоящий из 3 полипептидов: 48, 58, 93 кД. Стрихнин и глицин-связывающий сайт расположены на пептиде с Мг - 48 кД, который обладает способностью взаимодействовать с экзогенными лектинами. Встроенный в липосомы белок активирует транспорт ионов ОТ, который блокируется в присутствии стрихнина.
Иммунохимический анализ пептидных компонентов глицинового рецептора с помощью моноклональных антител позволил обнаружить существование общих антигенных детерминант этих рецепторных белков, выделенных из разных объектов: головного и спинного мозга мышей, крыс, свиньи и человека. Более того, интересными являются данные о том, что некоторые участки глицинового и ГАМК-рецепторов иммунологиче-ски идентичны. Этот факт хорошо подтвержден генно-инженерными исследованиями.
До недавнего времени предположение о существовании гомологии между нейрорецепторами I класса, т.е. быстродействующих инотропных рецепторов, выдвигалось лишь в качестве гипотезы. В последние годы одновременно в нескольких лабораториях было показано, что гены рецепторов ГАМК и глицина имеют гомологичные последовательности. Так, оказалось, что имеется примерно 50%-ная гомология между аминокислотными последовательностями а-субъединичной структуры глицинового рецептора с Мг = 48 кД и а- и р-субъединицами ГАМКА-рецептора. Обнаружена 25%-ная гомология между нуклеотидными последовательностями всех трех субъединиц н-ХР.
На рис. 9 представлены консервативные участки известных нейрорецепторов, выявленных при анализе нуклеотидных последовательностей. Характерными особенностями являются высокая степень в гомологии аминокислотной последовательности и расположении трансмембранных участков М1-М4. Обязательное присутствие двух цистеинов в районе 140-150 аминокислоты на расстоянии 14 нуклеотидов друг от друга — отличительная черта нейрорецепторов 1-го класса. Возможно, что все эти нейрорецепторы принадлежат одному семейству белков, кодируемых родственными генами.

Глутаматные рецепторы. Наличие глутаматсвязывающей активности, независимой от присутствия в среде ионов Na, обнаружено практически во всех структурах головного мозга. Наибольшее количество этих участков — в коре больших полушарий, гиппокампе, полосатом теле, среднем мозге и гипоталамусе.
Согласно современным представлениям, существует несколько подтипов глутаматных рецепторов. Их классифицируют прежде всего на основе изучения действия широко известных аналогов глутамата: N-Memn-D-аспартата, а-амино-З-пид-рокси-5-метил-4-изоксазол-пропионовой кислоты, каиновой кислоты, квискваловой кислоты. В табл. 5 представлена структура дикарбоновых возбуждающих аминокислот и некоторых их аналогов. В литературе принято выделять прежде всего два главных подтипа глутаматных рецепторов: NMDA- и не-NMDA-рецепторы. К He-NMDA-рецепторам относятся рецепторы АМРА и каиновой кислоты, сходные по своим физико-химическим свойствам и распространенности в структурах мозга.
Рассмотрим прежде всего NMDA-рецепторы. Они образуют довольно широко распространенный подтип рецепторов глутамата, которые участвуют в разнообразных событиях в ЦНС. В мозге млекопитающих NMDA-связывающие участки локализованы главным образом в кортикальных структурах, базальных ганглиях и сенсорно-ассоциативных системах; наивысшая их плотность обнаружена в гиппокампе. Считают, что они имеют отношение к целому ряду процессов возбуждения, формированию нейрональной пластичности и механизмам памяти, а также к патологическим явлениям нейрональной дегенерации в случае болезни Альцгеймера, церебральной ишемии и др.
Таблица 5
Структура кислых возбуждающих аминокислот и некоторых
их аналогов

NMDA-рецепторы состоят из ряда субъединиц сМг = 40-92 кД и легко олигомеризуются, образуя высокомолекулярные комплексы сМг = 230-270 кД. Эти белки являются гликопротеид-липидными комплексами, формирующими ионные каналы для катионов Na+, К+, Са+. Молекула глугаматного рецептора содержит большое количество гидрофобных аминокислот, которые связаны и с внутренней, и с внешней частью мембраны, организуя взаимодействие с липидами.
Рецептор NMDA имеет несколько участков, взаимодействующих аллостерически. Выделяют пять функционально различных участков, взаимодействие с которыми приводит к изменению активности рецептора:
1) участок связывания нейромедиатора;
2) регуляторный, или коактивирующий, глициновый участок;
3) участок внутри канала, который связывает фенциклидин и родственные соединения;
4) потенциал-зависимый Mg+- связывающий участок;
5) тормозной участок связывания двухвалентных катионов. Наиболее специфический синтетический агонист этих рецепторов — NMDA — не обнаружен в мозге. Предполагается, что кроме глутамата эндогенными медиаторами в этих рецепторах является L-аспартат и L-гомоцистеинат.
Из наиболее известных антагонистов рецепторов NMDA типа можно назвать 0-2-амино-5-фосфоновалериат' и D-2- ами-но-7-фосфоногептаноат. Более специфичны, однако, новые синтетические антагонисты: 3--пропил-Ь-фосфонат и МК-801.СРРл МК-801 - это неконкурентные ингибиторы NMDA, они не действуют непосредственно на участки связывания глутамата.
Своеобразна роль глицинового участка. Глицин в концентрации ОД мкМ увеличивает ответы NMDA-рецептора, и этот эффект не может быть заблокирован стрихнином /напомним, что последний является блокатором самостоятельных глициновых рецепторов). Сам глицин не вызывает ^ответа, а лишь увеличивает частоту открывания канала, не влияя на амплитуду тока при действии агонистов NMDA. Наличке глицина вообще необходимо, поскольку при полном его отсутствии рецептор не активируется L-глутаматом.
Самой важной функцией, которую осуществляет рецептор NMDA в ЦНС, является управление ионным каналом. Важным свойством является способность канала после связывания аго-ниста пропускать ионы Na+ и К+, а также ионы Са+. Предполагают, что внутриклеточный Са+, концентрация которого возрастает при участии рецепторов NMDA, вовлечен в инициацию процессов пластичности развивающегося и взрослого мозга.
Наибольшие токи при активации агонистами возникают при умеренной деполяризации мембраны: от -30 до -20 мВ и уменьшаются при высокой гиперполяризации или деполяризации; следовательно, ионные каналы NMDA-рецепторов являются в определенной мере потенциалзависимыми. Ионы Mg+ селективно блокируют активность рецепторов при таких сдвигах потенциалов. Ионы цинка также ингибируют ответ, но не имеют потенциалзависимого действия, очевидно влияя на другой участок связывания.
К другому подтипу рецепторов глутамата — не NMDA-pe-цепторам — относятся, в частности, рецепторы квискваловой кислоты. Изучение последних привело к пересмотру представления о том, что действие глутамата как нейромедиатора сводится лишь к деполяризации мембраны. Многие типы глутамат-ных рецепторов, и в особенности рецепторы квисквалата, могут функционировать как медленнодействующие метаботропные.
Они вполне соответствуют общим характеристикам метабо-тропных рецепторов, изложенным выше. Пептидная цепочка, составляющая их основу, содержит от 870 до 1000 аминокислотных остатков.
Часть He-NMDA-рецепторов—mGlnRl—реализует сигнал через О0-белки и систему внутриклеточных посредников: инози-толтрифосфатов, диацилглицерола, ионов кальция и др.
Другая разновидность метаботропных He-NMDA-рецепторов — mGlnR2 — реализует сигнал, подавляя синтез цАМФ или активируя синтез цГМФ.
Имеются сведения о том, что рецепторы этой категории участвуют в механизмах синаптогенеза и в изменениях, возникающих при деафферентации. В целом этот тип глутаматных рецепторов, как полагают, участвует в механизмах пластичности аналогично рецепторам NMDA. Но при этом активация рецепторов NMDA блокирует механизм инозитолфосфатной регуляции, связанной с He-NMDA-рецепторами, и наоборот: антагонисты NMDA усиливают действие глутамата на не-NMDA-pe-цепторы.
Весьма интересным примером современных методов изучения рецепторов служит цикл работ с кДНК и ^РНК, кодирующими белки глутаматных рецепторов. Существуют библиотеки полноразмерных генов или их фрагментов мозга млекопитающих. Имея поликлональные антитела к самым разнообразным нейрорецепторам, можно выделить с помощью иммунологического скрининга клоны ДНК, способные продуцировать искомые белковые фракции. Так, недавно из библиотеки кДНК были выделены клоны рекомбинантного фага, дающие положительный иммунологический сигнал на антитела, полученные к глутаматсвязывающему мембранному белку с Мг = 60 кД. Анализ ДНК, выделенной из этого фага, позволил обнаружить наличие вставки кДНК размером 500 нлт, которая способна продуцировать белок с Мг = 14 кД и соответствует узнающей субъединице глутаматного рецептора.-^С помощью этой ДНК была выделена фракция мРНК, комплементарная данной последовательности ДНК. Для доказательства, что выделенная фракция мРНК кодирует синтез глутаматных рецепторов, она была инъецирована в ооциты лягушки, которые являются удобным объектом изучения электрофизиологических свойств нейрорецепторов. Ооциты лягушки обладают эффективным белок-синтезирующим аппаратом, но не имеют собственных нейрорецепторов. После инъекции чужеродной мРНК был измерен мембранный потенциал ооцитов в присутствии глутамата и его аналогов. Оказалось, что выделенная фракция мРНК способна кодировать синтез de novo глутаматных рецепторов каинатного типа.
Возможность одновременного синтеза всех подтипов глутаматных рецепторов в ооцитах лягушки была продемонстрирована другими исследователями. Введение тотальной мРНК, выделенной из мозга крыс, приводило к появлению электрофизиологических ответов у ооцитов на аппликацию NMDA, каината и квисквалата. Более того, ионные токи, регистрируемые на мембране, мало отличались от таковвгх:, обнаруженных на мембранах нейронов. Были, таким офазом, представлены убедительные факты в пользу того, что основные компоненты рецепторного комплекса для глутамата синтезируются совместно, причем биосинтез их не зависит от клетки-носителя и типа мембраны, в которую они затем встраиваются.
Перспективными являются исследования шклада глутаматных рецепторов в патохимию ряда заболеваний ЦНС. Полагают, что эти нейрорецепторы могут служить маркерами деструктив
ных повреждений возбуждающих глутаматергических путей головного мозга и участвовать в аутоиммунных реакциях организма человека. Установление роли глутаматных рецепторов в патогенезе нервно-психических заболеваний — это не единственное направление современной медицины. Появились уже конкретные примеры использования разных антагонистов глутаматных рецепторов против явлений укачивания, токсического действия высоких парциальных давлений кислорода, при лечении инсультов и др.. Кроме того, антагонисты глутаматных рецепторов могут составить основу для создания малотоксичных инсектицидных препаратов для сельского хозяйства.
Адренорецепторы История изучения адренорецепторов тесно связана с открытием биологической функции катехолами-нов в клетках надпочечников. Гипотеза о существовании этого вида рецепторов в самых разнообразных клетках наряду с исследованиями ХР оказалась наиболее плодотворной для развития теории взаимодействия физиологически активных веществ с рецепторами. Несмотря на то что адренорецепторы ъ нервной ткани присутствуют в относительно небольшом количестве, они играют важную роль в регуляции психоэмоциональных функций и деятельности всех отделов сердечно-сосудистой системы.
Адренорецепторы
подразделяют на два типа:![]() , в
зависимости от связывания адренотропных лигандов. Адренорецепторы
, в
зависимости от связывания адренотропных лигандов. Адренорецепторы![]() и
и![]() , в свою очередь, могут
быть разделены на подклассы
, в свою очередь, могут
быть разделены на подклассы ![]() и
и![]() ,
,![]() и
и![]()
Для![]() ,
,![]() ,
,![]() и
и![]() -рецепторов характерна
преимущественно постсинаптическая локализация, а для а2-,, вторичный
мессенд-жер, служащий активатором опять-таки протеинкиназ, но другого типа,
нежели протеинкиназы, активируемые в результате включения aj-рецепторов.
-рецепторов характерна
преимущественно постсинаптическая локализация, а для а2-,, вторичный
мессенд-жер, служащий активатором опять-таки протеинкиназ, но другого типа,
нежели протеинкиназы, активируемые в результате включения aj-рецепторов.
G-белки — передатчики и модификаторы сигнала от собственного рецепторного белка — также имеют существенные отличия в разных классах адренергических рецепторов. Важным общим их свойством является взаимодействие х ГТФ и ГДФ. В неактивном состоянии они связаны с ГДФ. При взаимодействии нейромедиатора с рецепторным белком последний вступает в контакт с G-белком и меняет его конформацию так, что на место ГДФ становится ГТФ. С этого момента G-белок приобретает способность воздействовать на активность белка-эффектора. Однако G-белок является ГТФазой, быспро расщепляющей ГТФ до ГДФ. В результате он переходит в исходное состояние.
Заметим,
что G-белок
а2-адренорецетхиэтав, обозначаемый Gs,
активирует аденилатциклазу, обеспечивая, >в конечном счете, повышение уровня
цАМФ и, далее, стимуляцию протеин-киназ. G-белки![]() - и
- и![]() -адренорецепторов — G-белки
обеспечивают, напротив, подавление активности щиклазы, снижение уровня цАМФ и,
соответственно, снижение активности определенных протеинкиназ.
-адренорецепторов — G-белки
обеспечивают, напротив, подавление активности щиклазы, снижение уровня цАМФ и,
соответственно, снижение активности определенных протеинкиназ.
Собственно рецепторный белок представляет собой пептидную цепочку с Мг порядка 60-80 кД, пронизывающую внешнюю мембрану так, что ее С-конец и ряд петлеобразных участков экспонированы наружу, а N-конец и опять-таки ряд петлеобразных участков обращены внутрь клетки. Нейромедиатор, взаимодействуя с обращенными наружу участками цепи, меняет конформацию всего белка, так что участки, обращенные внутрь, приобретают сродство к Gg-белку, Gj-белку или к О0-белку. "Выбор" одного из этих белков определяется тонкими особенностями структуры R-бел-ка в области, обращенной внутрь клетки. Заметим также, что участки пептидной цепи, содержат по 20-25 преимущественно гидрофобных аминокислот, а петлевидные участки вне мембраны состоят преимущественно из гидрофильных аминокислот. N-конец полипептида обычно яддикозилирован, а на С-конце есть остатки серина и треонина, wropbie могут фосфорилироваться; последний процесс модулирует активность рецептора.
Практически
любое изменение функции![]() и
и![]() -адренорецепторов сопровождается
активацией системы внутриклеточных посредников, которые способны избирательно
передавать внешний сигнал в цитоплазму и на генетический аппарат клетки. В этом
случае геном нейрона может регулировать биосинтез мембранных компонентов или
активировать процессы, связанные с их фосфорилированием. Последние реакции
приводят к изменению хемочувствительности нервных клеток, иными словами,
изменению "информационной емкости" нейронов, и их связывают с
механизмом запоминания.
-адренорецепторов сопровождается
активацией системы внутриклеточных посредников, которые способны избирательно
передавать внешний сигнал в цитоплазму и на генетический аппарат клетки. В этом
случае геном нейрона может регулировать биосинтез мембранных компонентов или
активировать процессы, связанные с их фосфорилированием. Последние реакции
приводят к изменению хемочувствительности нервных клеток, иными словами,
изменению "информационной емкости" нейронов, и их связывают с
механизмом запоминания.
Дофаминовые рецепторы. Прогресс в изучении структуры и функции дофаминовых рецепторов прежде всего был связан с обнаружением их антагонистов: производных фенотиазепама, галоперидола, спиропервдола и др. Радиолигандные исследования с помощью этих соединений показали, что в головном мозге имеются участки специфического связывания Н-галоперидола, Н-спироперидола и Н-апоморфина с параметрами, колеблющимися в пределах Кд ~7 нМ и Вмакс — 240-400 пмоль/мг белка.
Гетерогенность дофаминовых рецепторов ЦНС подтверждается не только биохимическими и фармакологическими экспериментами in vitro. Анализ влияния различных дофаминсодер-жащих препаратов на поведение крыс обнаружил разные стереотипные двигательные реакции. Оказалось, что вращение крыс в ту или иную сторону опосредуется двумя разными классами дофаминовых рецепторов.
Все дофаминергические рецепторы являются метаботропны-ми, сопряженными с аденилатциклазой. Они классифицируются на 4 типа: Д^, Д2-, Д3 - и Д4 по параметрам связывания их с агонистами и антагонистами и по белковой системе, трансформирующей сигнал. Наиболее изучены Др и Д2-рецепторы. Д j -рецепторы активируют аденилатциклазу через 6£-белок, а Д2-рецепторы, напротив, подавляют аденилатциклазу через Gs-белок. Первые подавляются лишь относительно высокими, микромолярными концентрациями нейролептиков, в то время как для подавления вторых достаточны наномолярные концентрации. Количество и распространение Дри Д2-рецепторов выше, чем Д3 и Д4 в ткани мозга, Д2-рецепторы локализованы преимущественно в клетках ниг-ростриарной и мезкортиколимбической систем. Рвотное действие апоморфина — результат его взаимодействия с Д2-рецепто-рами. Именно нарушение, несбалансированное усиление деятельности систем, связанных с Д2-рецепторами, ассоциируют с синдромом шизофрении. Подавление Дг- и Д2-рецепторов индуцирует нарушения различных форм стереотипных двигательных реакций. Одно из их проявлений — болезнь Паркинсона.
Рецепторы серотонина. Описание рецепторов серотонина облегчается их сходством с рецепторами норадреналина и дофамина. Это метаботропные рецепторы, локализованные как в мозге, так и на периферии. Известно несколько подтипов рецепторов серотонина, в частности 5НТ\, сопряженный с аде-нилатциклазой, и 5НТ2, сопряженный с фосфолипазой С. Последний особенно распространен в постсинаптических мембранах мозга. Специфические антагонисты — спиперон и кетансерин. Сложным является отношение этих рецепторов с производными лизергиновой кислоты, в том числе LSD. По отношению к некоторым разновидностям 5НТ2-рецепторов он является агонистом, а к другим — антагонистом.
Активация 5НТ2 входит в систему реакций, индуцирующих ортодоксальный сон. По-видимому, дефицит или ингибирова-ние 5НТ2 связано с депрессивными состояниями, а активация некоторых из них — с галлюцинациями. Нарушения серотонин-ергической системы связывают и с такой патологией, как тяжелые формы мигрени. Существует характерный двигательный синдром, регулируемый 5НТ2-рецепторами, "отряхивание мокрой собаки", используемый нередко в исследованиях серотонинергической системы.
Функции 5НТ2-рецепторов, также представленных и в мозге, и на периферии, изучены в меньшей мере. Одна из причин этого — отсутствие высокоспецифичных антагонистов.
Многообразными являются периферические эффекты активации рецепторов серотонина всех подтипов. Главные из них состоят в сокращении сосудов, положительных хроно- и инотропных воздействиях на сердце, а также в стимуляции аггрегации тромбоцитов.
Рецепторы гистамина. К настоящему времени выявлены и исследованы три типа рецепторов гистамина: Hj и Н? — постсинаптические и Н3 - пресинаптические. Рецепторы Hi являются метаботропными, они сопряжены с фосфолипазой С и индуцируют образование инозитол-трифосфата и диацилглицерола. Ингибиторами Я^рецепторов служат димедрол, фенкарол, супрастин и другие соединения, получившие широкое применение в медицине. Метаботропными являются также рецепторы Н2, однако они сопряжены с аденилатциклазой и индуцируют повышение уровня циклического АМФ. Хорошо изученным ингибитором Н2-рецепторов является циметидин известный противоязвенный агент. Пресинаптические рецепторы гистамина Н3 являются ауторецепторами. Они подавляют выход гистамина из нервных окончаний, т.е. осуществляют обратную связь между уровнем гистамина в синапсе и его секрецией.
Рецепторы пуринов. Последнее десятилетие ознаменовалось накоплением данных об обширном семействе рецепторов разнообразных пуринов: аденозина, АТФ, относительно мало изученного макроэргического соединения диаденозинтетрафосфа-та, а также АМФ и АДФ. Рецепторы пуринов делят на две большие группы: рецепторы, преимущественно взаимодействующие с аденозином, и рецепторы, преимущественно взаимодействующие с АТФ и АФА. Рецепторы аденозина Р1 являются медленными метаботропными рецепторами. Среди них есть рецепторы, подавляющие системы синтеза цАМФ, и рецепторы, напротив, активирующие синтез цАМФ.
Особого рассмотрения заслуживают свойства рецепторов А1. Значительная их часть локализована на пресинаптических мембранах. Довольно специфическими ингибиторами рецепторов А1 являются теофилин и кофеин. Иначе говоря, введение этих широко известных психостимулирующих агентов ведет к увеличению уровня цАМФ в некоторых постсинаптических структурах. Заметим, что кофеин и теофилин являются также довольно специфичными ингибиторами фосфодиэстераз, что тоже способствует повышению уровня цАМФ. Такое двустороннее действие этих соединений на уровень цАМФ позволяет понять их особое значение как психостимуляторов.
Рецепторы А1 включаются не только аденозином, но и АМФ, причем последний может происходить не только из нервных окончаний, но и из других образований мозга. Истощение энергетических систем мозга, связанное с образованием АМФ, может служить, таким образом, сигналом для включения рецепторов А1. Это позволяет лучше понять ряд физиологических эффектов, которые наблюдаются при срабатывании рецепторов А1: успокаивающие, седативные, противосудорожные. Иначе говоря, эти рецепторы выступают как защитники энергетических резервов мозга в экстремальных ситуациях. С этим же сопряжены их гипотензивные эффекты. Из периферических реакций на включение рецептора А1 отметим брадикардию и воздействие на автономные проводящие системы сердца. Адено-зин и его производные оказались перспективными средствами для лечения аритмий.
Из общих физиологических эффектов аденозиновых рецепторов А2 отметим стимуляцию глюконеогенеза и подавление агрегации тромбоцитов.
Рецепторы Р2, имеющие наибольшее сродство к АТФ и АФ4А и наименьшее к аденозину и АМФ, также делятся на несколько подтипов. В отличие от аденозиновых рецепторов среди них есть быстрые, канальные рецепторы заслуживает периферический эффект такого макроэргического соединения, как АДФ —стимуляция тромбоцитов.
Рецепторы нейропептидов. Все рецепторы нейропептидов являются метаботропными, медленными рецепторами. Наиболее изучены опиатные рецепторы.
Опиаты морфин и родственные ему соединения — известны в медицине с древних времен. Их применяли, как правило, в качестве обезболивающих и наркотических средств, способных влиять на психику человека. Однако хэпиатные рецепторы, которые являются объектами действия опиатов в ЦНС, открыты сравнительно недавно, в начале 70-х годов. Фармакологическими исследованиями было показано, что ОР связывают большое число синтетических и природных лигандов. Первыми изученными лигандами ОР были экзогенные вещества — морфин, дигидроморфин, норморфин, леворфанол и др. Из алкалоидов растительного происхождения наиболее известны налоксон, пентазоцин и налтрексон. Два последних обладают свойствами частичных антагонистов. Эндогенные пептиды — эндорфины, энкефалины и динорфины —взаимодействуют аналогично морфину и относятся к агонистам ОР.
ОР гетерогенны по составу, и классифицируют их на основе взаимодействия со специфическими лигандами. Они подразделяются на субтипы: мю-, дельта-, каппа-, сигма- и эпсилон-рецепторы. Принято считать, что морфин выявляет мю-тип рецепторов, Ала, Лей-энкефалин, дельторфины и некоторые другие аналоги энкефалинов — дельта-рецепторы, кетоциклазоцин и динорфины —каппа-рецепторы, N-аллилнорияклазоцин — сигма-рецепторы, а р-эндорфин мю- и эпсилон-рецепторы. Вместе с тем следует помнить, что эти лиганды не обладают абсолютной специфичностью и могут частично взаимодействовать с разными субтипами О Р.
Связывание радиолигандов свидетельствует о различном распределении субтипов ОР по структурам головного мозга.
Для изучения биохимических характеристик опиатных рецепторов многие годы предпринимались попытки выделить их в нативном состоянии из биологических мембран мозга. Однако ОР оказались трудным объектом, они инактивировались под действием ионных детергентов, которые ранее успешно применялись для солюбилизации других рецепторов. Кроме того, попытка полного удаления липидного окружения из препаратов ОР также вела к инактивации.
Лишь использование неионных детергентов типа дигитонинаглиоксихолата и CHAPS — позволяет солюбилизировать из мембран белки рецепторного комплекса с сохранением 50-70% опиатсвязывающей активности, ОР-рецепторы оказались ли-попротеидами. Роль липидных компонентов и окружения ОР в связывании опиоидов пока не ясна, однако, присутствие сульфоцереброзидов существенно увеличивает уровень связывания агонистов с рецепторами, и было сделано предположение о том, что сульфоцереброзиды входят в активный центр ОР.
Собственно рецепторные лиганд-связывающие компоненты ОР имеют молекулярную массу порядка 45000-66000 дальтон. В целом ОР подобны другим метаботропным рецепторам. Взаимодействие с лигандом ведет к включению G-белков и, в конечном счеге, к подавлению аденилатциклазы. Интересно, что молекула рецептора обладает дефосфорилирующей активностью. Она усиливается теми же факторами, которые усиливают связывание опиатов. При этом дефосфорилирование изменяет ли-ганд-рецепторное взаимодействие.
Известно также, что активация опиатных рецепторов сопровождается повышением К+ проводимости мембраны и/или снижением Са+ проводимости мембраны, что ведет к принципиально общему конечному результату — уменьшению входа Са+ в пресинаптические окончания во время прохождения потенциала действия и к снижению количества высвобождаемого нейромедиатора или гиперполяризации постсинаптической мембраны.
Ферменты-рецепторы. Название этого типа рецепторов вызывает некоторое недоумение, ибо конечный компонент мета-ботропных рецепторов является, как правило, ферментом, например циклазой или фосфолипазой. Однако последние служат лишь компонентом комплекса белков, составляющих метаботропный рецептор. Медиатор при этом не взаимодействует непосредственно с ферментом. Существует, однако, ряд систем межнейрональной передачи сигнала, когда медиатор прямо действует на фермент. Приведем лишь один пример. Относительно новый нейромедиатор нитроксид прямо активирует гуанилатциклазу и синтез цГМФ.
Выводы
1. Обязательным звеном передачи нервного импульса в химических синапсах являются рецепторы — образования, состоящие из белков и гликолипидных компонентов, которые с высокой специфичностью связывают нейромедиатор, меняют конформацию и обеспечивают трансформацию сигнала в изменения ионных потоков через мембрану и в образование вторичных мессенджеров в клетке.
2. По типу вызываемых медиатором процессов рецепторы делятся на две категории: 1) быстродействующие, содержащие в своей структуре ионный канал, открытие которого ведет к изменению потенциала мембраны; 2) медленнодействующие, состоящие из компонентов, периодически связывающихся друг с другом, которые после взаимодействия с нейромедиатором запускают цепь реакций, образующих вторичные молекулы — посредники, циклические нуклеотиды, диацилглицерол, ино-зитолфосфаты и др.
3. Одни и те же медиаторы в разных синапсах могут взаимодействовать с рецепторами разных типов и разных подтипов.
4. Современный уровень понимания структуры и механизма действия многих рецепторов таков, что оказалось возможным "собирать" активные рецепторы из компонентов вне клеток — в мембранах липосом — и даже индуцировать с помощью соответствующих мРНК их синтез, встраивание в мембраны и действие в клетках, не содержащих ранее этих образований.
5. Действие ряда важнейших фармакологических агентов, используемых при лечении заболеваний ЦНС, направлено на рецепторы нейромедиаторов — их активацию или подавление.
© 2009 База Рефератов