
Рефераты по рекламе
Рефераты по физике
Рефераты по философии
Рефераты по финансам
Рефераты по химии
Рефераты по хозяйственному праву
Рефераты по цифровым устройствам
Рефераты по экологическому праву
Рефераты по экономико-математическому моделированию
Рефераты по экономической географии
Рефераты по экономической теории
Рефераты по этике
Рефераты по юриспруденции
Рефераты по языковедению
Рефераты по юридическим наукам
Рефераты по истории
Рефераты по компьютерным наукам
Рефераты по медицинским наукам
Рефераты по финансовым наукам
Рефераты по управленческим наукам
психология педагогика
Промышленность производство
Биология и химия
Языкознание филология
Издательское дело и полиграфия
Рефераты по краеведению и этнографии
Рефераты по религии и мифологии
Рефераты по медицине
Реферат: Регенерация периферической нервной системы позвоночных
Реферат: Регенерация периферической нервной системы позвоночных
РЕГЕНЕРАЦИЯ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ
Восстановление поврежденных аксонов
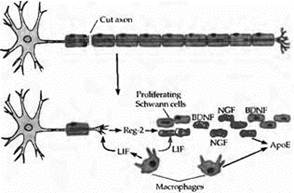
Шванновские клетки периферической нервной системы обеспечивают окружение, способствующее регенерации аксона. Стимулирующая рост нейронов активность шванновских клеток является следствием секреции многих трофических факторов, экспрессии на поверхности клеток молекул адгезии и интегринов, а также продукции компонентов внеклеточного матрикса, таких как ламинин. Например, эксперименты, в которых повреждался седалищный нерв, показали, что в то время, как периферический участок аксона дегенерирует, уцелевшие шванновские клетки в этой области синтезируют в высоких концентрациях два нейротрофических фактора: BDNF (нейротрофический фактор, полученный из мозга) и NGF. Таким образом, шванновские клетки могут снабжать трофическими факторами BDNF и NGF двигательные, чувствительные и симпатические аксоны, регенерирующие к своим периферическим мишеням. Интересно, что такие «денервированные» шванновские клетки экспрессируют на своей поверхности большое число низкоаффинных рецепторов для NGF и BDNF. Возможно, это поддерживает некоторую тоническую концентрацию этих нейротрофинов, которые служат для выбора правильного направлении для роста регенерирующих аксонов84). После регенерации нервов шванновские клетки прекращают продукцию NGF и BDNF и вновь обеспечивают изоляцию аксонов.
Аполипопротеин E (ApoE), синтезируемый шванновскими клетками и макрофагами, также накапливается в дистальном участке поврежденных периферических нервов и связывается с базальной мембраной шванновских клеток (см. рис. 2 ). АроЕ поддерживает жизнеспособность нейронов за счет защиты клеток от окислительного повреждения, а также стимуляции роста и адгезии клеток.
|
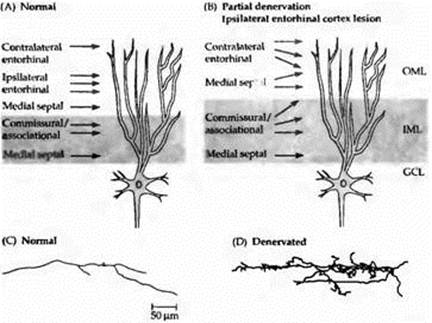
Рис.1. Спраутинг аксонов в ЦНС млекопитающих. (А) Типичная гранулярная клетка зубчатой фасции на ее дендритах во внешнем молекулярном слое (OML) получает множественные сина птические входы от ипсилатеральной энторинальной коры головного мозга и единичные входы от контралатеральной коры и медиальной перегородки. Дендриты внутреннего молекулярного слоя (IML) получают входы от медиальной перегородки и коммиссуральных/ассоциативных волокон. GCL= гранулярный клеточный слой. (В) Удаление ипсилатеральной энторинальной коры приводит к интенсивному росту аксонов нервных клеток энторинальной коры с контралатеральной стороны и замещению ипсилатерального энторинального входа в OML. Наблюдается также спраутинг аксонов, приходящих в DML от медиальной перегородки. Коммиссуральные/ассоциативные волокна в IML расширяют зону иннервации. (С, D) Примеры терминальных разветвлений аксонов от контралатеральной энторинальной коры в OML зубчатой фасции. (С) Норма. (D) Два месяца спустя после повреждения ипсилатеральной энторинальной коры. |
Тем не менее, у нокаутных мышей, лишенных АроЕ, регенерация и ремиелинизация периферических аксонов существенно не меняется. Однако мыши с отсутствием АроЕ имеют сниженное число немиелинизированных аксонов и, соответственно, сниженную чувствительность к повреждающим тепловым стимулам.
При повреждении периферического нерва в зоне повреждения активируются факторы, стимулирующие пролиферацию шванновских клеток. Они включают два цитокина: фактор, ингибирующий лейкемию (ФИЛ), и Reg-2, являющийся потенциальным митогеном шванновских клеток (см. рис. 2). Reg-2 специфически экспрессируется в развивающихся и регенерирующих двигательных и чувствительных нейронах, и эта экспрессия усиливается ФИЛ. Антитела к Reg-2 замедляют регенерацию после передавливания седалищного нерва.
Специфичность реиннервации
Для полного восстановления функции после повреждения нервной системы регенерирующие аксоны должны восстановить связи со своими прежними мишенями. Классические эксперименты Лэнгли, подтвержденные позже исследованиями одиночных клеток, показали, что регенерирующие преганглионарные аксоны нейронов автономной нервной системы реиннервируют соответствующие постганглионарные нейроны. В этом механизме имеет значение, по-видимому, сигнал о позиции нейрона, который оказывает влияние на формирование синапса между нейроном и клеткой-мишенью.
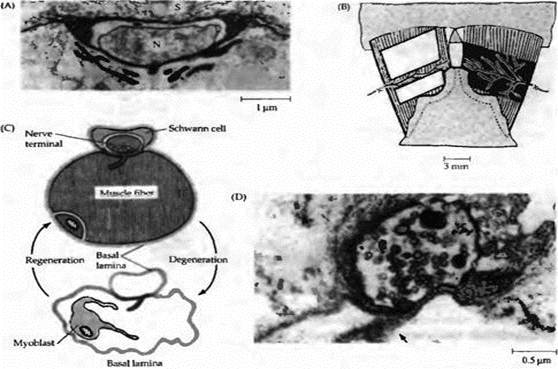
| Рис. 2. Шванновские клетки способствуют регенерации аксонов в периферической нервной системе позвоночных. После аксотомии дистальный участок аксона и миелиновая оболочка дегенерируют и фагоцитируются. Пролиферация шванновских клеток стимулируется двумя цитокинами: фактором, ингибирующим лейкемию (LIF), из макрофагов и Reg-2 из окончаний аксонов. Экспрессия Reg-2 усиливается LIF. Пролиферирующие шваннов ские клетки синтезируют два нейротрофических фактора, BDNF и NGF, которые сохраняются на клеточной поверхности их низкоаффинными рецепторами. Нейротрофины поддерживают процесс регенерации аксонов и направляют их к соответствующим мишеням. Шванновские клетки и макрофаги синтезируют также аполипопротеин Ε (ΑροΕ), который способствует выживанию нейронов и регенерации их аксонов. |
|
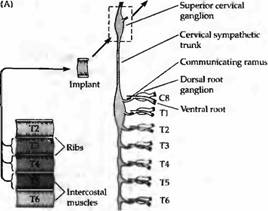
Этот механизм справедлив как для симпатической, так и двигательной системы. Так, если межреберная мышца, извлеченная из передней части грудной клетки, трансплантируется в область шеи, то она преимущественно реиннервируется аксонами шейного симпатического ствола, расположенными в самых передних участках спинного мозга (рис. 3). Наоборот, мышцы, трансплантированные из задних областей тела, имеют тенденцию к реиннервации аксонами более дорзальньгх нейронов. Подобная тенденция установлена и для трансплантации симпатических ганглиев различного исходного расположения. У молодых крыс зависимость от позиции нейрона показана в реиннервации мышц, имеющих множественную иннервацию несколькими сегментами.
Нервно-мышечные синапсы у новорожденных крыс, головастиков и взрослых тритонов после поперечного пересечения двигательного нерва с высокой точностью воссоединяются с исходной синаптической зоной . Одним из механизмов избирательного восстановления синапсов является конкуренция между аксонами. В мышцах саламандры, иннервированных чужим аксоном, чужеродный синапс элиминируется после того, как нормальный нерв восстановил свой синаптический контакт. У взрослых млекопитающих чувствительные, двигательные и постганглионарные аксоны показывают более низкую избирательность по отношению к их исходным мишеням. Чужие нервы при иннервации мышечных волокон могут быть столь же эффективными, как и оригинальные нервные волокна. Более того, чужой нерв может даже вытеснить исходный аксон в интактных мышцах взрослой крысы). Избирательная регенерация у взрослых млекопитающих возможна, если периферический нерв был не перерезан, а передавлен так, чтобы сохранились эндоневрий и базальная мембрана шванновских клеток, окружавших интактные аксоны (см. рис. 4)). При таких условиях регенерирующие аксоны направляются вновь к своим исходным мишеням. Если эндоневрий был разрушен, например при перерезке нерва, регенерация происходит беспорядочно и аксоны часто создают синапсы с неподходящими мишенями.
|
|
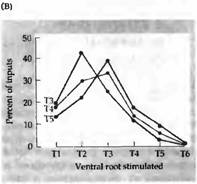
| Рис. 3. Селективная реиннервация мышц, основанная на сегментарном происхождении. (А) Межреберная мышца из грудного сегмента ТЗ, Т4 или Т5 была трансплантирована на шею взрослой крысы и после удаления верхнего шейного ганглия была реиннервирована преганглионарными аксонами шейного симпатического ствола. Сегментарное происхождение входов было определено регистрацией активности мышц при стимуляции отдельных вентральных корешков, которые соединяют преганглиальные аксоны с нервным стволом. (В) Распределение входов к трансплантированным ТЗ, Т4 и Т5 мышцам. Передние мышцы склонны к реиннервации передними преганглиальными аксонами. |
Свойства нерва и мышцы после образования синапса чужим нервом
Первые наблюдения о последствиях формирования синапсов чужим нервом относятся к 1904 году, когда Лэнгли и Андерсон провели выдающееся исследование, показавшее, что мышцы кошки могут иннервироваться преганглионарными холинергическими симпатическими волокнами, которые в норме создают синапсы в автономном ганглии . Было также показано формирование подобных синапсов между автономными нервами лягушки и скелетными мышцами крысы (рис. 4.). Многие свойства нерва и мышцы в таких экспериментах оставались без изменений, несмотря на необычную иннервацию.
В других экспериментах было показано, что свойства мышц с чужеродной иннервацией заметно изменяются. Ярким примером являются медленные скелетные мышечные волокна лягушки: они иннервированы диффузно, имеют характерные детали строения и не генерируют регенеративных потенциалов действия или быстрых мышечных сокращений. После денервации медленные волокна могут быть реиннервированы нервами, которые в норме иннервируют быстрые мышцы. В этих условиях медленные волокна становятся способными генерировать потенциалы действия и осуществлять быстрое мышечное сокращение. Экклс и коллеги после перерезки меняли местами нервы, иннервирующие два типа мышц котят и крыс, различающихся по скорости сокращения. Оба этих типа мышечных волокон генерируют распространяющиеся потенциалы действия и называются медленно- и быстро-сокращающимися волокнами, соответственно. После реиннервации чужим нервом медленно-сокращающиеся мышцы становились быстрее, а быстро-сокращающиеся — медленнее. Главным фактором трансформации являлся паттерн импульсов в нерве и результирующие мышечные сокращения; мотонейроны, иннервирующие медленно- и быстро-сокращающиеся мышечные волокна, возбуждаются с различной частотой Роль базальной мембраны в регенерации нервно-мышечных синапсов
Структурой, играющей ключевую роль в регенерации нервно-мышечных синапсов, является синаптическая базальная мембрана, которая находится между нервным окончанием и мышечной мембраной. Синаптическая базальная мембрана представляет собой интенсивно окрашиваемое внеклеточное вещество, состоящее из протеогликанов и гликопротеинов. Как показано на рис. 3А, базальная мембрана окружает мышцу, нервное окончание и шванновскую клетку и погружается в складки постсинаптической мембраны.
МакМахан и его коллеги провели серию элегантных исследований роли синаптической базальной мембраны в дифференцировке нерва и мышцы. Успех их работы базировался на использовании очень удобной экспериментальной модели тонкой кожно-грудной мышцы лягушки, в которой положение концевых пластинок легко различимо в живой мышце. На первом этапе клетки в определенном участке мышцы локально разрушали либо перерезкой нерва и мышечных волокон, либо повторным приложением металлической пластины, охлажденной жидким азотом (рис. 3В). В течение нескольких дней часть мышечных волокон в участке повреждения дегенерировала вместе с нервными окончаниями и была фагоцитирована, но базальная мембрана оставалась при этом интактной (рис. 3C). Место исходного нервно-мышечного соединения могло быть легко распознано по характерной морфологии базальной мембраны мышцы и шванновских клеток, остающихся в зоне синапса, а также по наличию холинэстеразы, сохраняющейся на синаптической базальной мембране и в синаптических складках.
Через две недели после повреждения в зоне синаптической базальной мембраны формировались новые мышечные волокна, контактирующие с регенерирующими аксональными окончаниями. Стимуляция нервов вызывала сокращения вновь образованных мышечных волокон. Почти все регенерированные синапсы локализовались точно на исходных синаптических зонах, что выявлялось после окраски мышцы на присутствие холинэстеразы. Таким образом, был сделан вывод о наличии сигналов, ассоциированных с синаптической базальной мембраной, которые определяют успех регенерации синапсов.
Синаптическая базальная мембрана и формирование синаптической специализации
Для дальнейшего исследования природы сигналов, ассоциированных с синаптической базальной мембраной, мышцы повреждали, нерв раздавливали, а регенерацию мышечного волокна предотвращали рентгеновским облучением. Регенерирующие аксоны росли к исходным синаптическим зонам, что выявлялось окраской на холинэстеразу, и формировали активные зоны для освобождения медиатора точно напротив участков базальной мембраны, связанных со вторичными синаптическими складками, — и все это происходило в отсутствие клеточных элементов постсинаптической мишени (рис.3D).
В параллельной серии экспериментов МакМахан и его коллеги продемонстрировали, что синаптическая базальная мембрана в регенерирующих мышечных волокнах содержит факторы, запускающие дифференцировку постсинаптической мембраны. Мышцы повреждались, как было описано ранее, а реиннервация предотвращалась удалением большого сегмента нерва. При регенерации новые мышечные волокна образовывали вторичные складки и кластеры АХ рецепторов и ацетилхолинэстеразы точно в зоне контакта с исходной синаптической базальной мембраной (рис. 4). Таким образом, сигналы, ассоцированные с синаптической базальной мембраной, при регенерации могут инициировать формирование синаптических специализаций как в мышечных волокнах, так и в нервных окончаниях.
Идентификация агрина
Для идентификации сигнала, связанного с базальной мембраной и инициирующего постсннаптическую дифференцировку, МакМахан и его коллеги использовали морского ската Torpedo califomica. Из электрических органов этого животного, родственных скелетной мышце, они приготовили экстракты, содержащие базальные мембраны. Добавленные к культуре мышечных волокон, экстракты симулировали эффекты синаптической базальной мембраны на регенерацию мышечных волокон, а именно индуцировали формирование кластеров АХ рецепторов вместе с другими компонентами постсинаптической мембраны (рис. 5).
Активный компонент экстрактов, названный агрином, был очищен и охарактеризован, а у цыпленка, крысы и ската клонирована соответствующая кДНК.
|
| Рис. 4. Базальная мембрана и регенерация синапсов. (А) Микрофотография нормального нервно-мышечного синапса лягушки, окрашенного рутением красным, показывающая базальную мембрану, погруженную в постсинаптические складки и окружающую шванновскую клетку (S) и нервное окончание (N). (В) Изображение кожно-грудной мышцы, показывающее замороженный (справа) или перерезанный (слева) участок, с целью вызвать локальное повреждение мышечных волокон. (С) Замораживание приводит к дегенерации и фагоцитозу всех клеточных элементов нервно-мышечного соединения, оставляя целой только базальную мембрану мышечного волокна и шванновской клетки. Новые нервно-мышечные синапсы создаются регенерирующими аксонами и мышечными волокнами. (D) Нерв и мышца были повреждены, регенерация мышечных волокон предупреждена рентгеновским облучением. В отсутствие мышечных волокон аксоны регенерировали, восстанавливали связь с исходными синаптическими зонами и формировали активные зоны. |
Результаты гибридизации in situ и иммуногистохимических исследований продемонстрировали, что агрин синтезируется мотонейронами, транспортируется по аксонам и, освобождаясь, индуцирует дифференцировку постсинаптического аппарата в развивающихся нервно-мышечных синапсах. Затем агрин становится частью синаптической базальной мембраны, где участвует в сохранении постсинаптического аппарата и запускает дифференцировку во время регенерации.
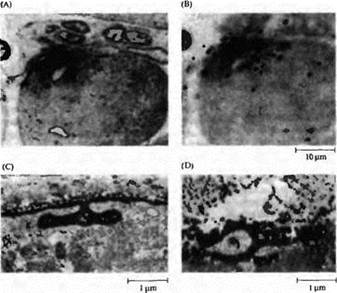
| Рис. 5. Аккумуляция AX рецепторов и ацетилхолинэстеразы в исходных синаптических зонах мышечных волокон, регенерирующих в отсутствие нерва. Мышца была заморожена, как на рис. 24.13В, но регенерация нерва была блокирована. Новые мышечные волокна сформировались в пределах оболочек базальной мембраны. (А и В) Ауторадиография регенерированной мышцы с окраской на холинэстераэу, для того чтобы выделить исходную синаптическую зону (в фокусе в части А), и помеченной радиоактивным -бунгаротоксином, чтобы определить положение АХ рецепторов (серебряные зерна в фокусе в части В). (С) Электронная микрофотография исходной синаптической зоны в регенерированной мышце, отмеченная пероксидазой хрена (HRP), конъюгированной с -бунгаротоксином. Распределение АХ рецепторов обнаруживается по высокой плотности продукта реакции с HRP, который позволяет различать поверхность мышечного волокна и синаптические складки. (D) Электронная микрофотография исходной синаптической зоны в регенерирующей мышце с окраской на холинэстеразу. |
Исходная холинэстераза деградировала после заморозки мышцы. Таким образом, выявляемый продукт реакции обусловлен холинэстеразой, синтезированной и аккумулированной в исходной синаптической зоне регенерирующим мышечным волокном. |
Специфичный для мышцы рецептор тирозинкиназы, называемый MuSK, образует часть агринового рецептора. Активация MuSK инициирует внутриклеточное фосфорилирование, что служит пусковым сигналом для агрегации АХ рецепторов.
Литература:
1. Sanes, J. R., and Lichtman, J.W. 1999. Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci. 22: 389-442.
2. Song, H-J., and Poo, M-M. 1999. Signal transduc-tion underlying growth cone guidance by diffusible factors. Curr. Opin. Neurobiol. 9: 355-363.
3. Walsh, F. S., and Doherty, P. 1997. Neural cell adhesion molecules of the immunoglobulin super-family: Role in axon growth and guidance. Annu Rev. Cell Dev. ВЫ. 13: 425-456.
4. Zigmond, M.J., Bloom, F. E., Landis, S.C., Roberts, J.L., and Squire, L. R. (eds.). 1999. Fundamental Neuroscience. Academic Press, New York.
© 2009 База Рефератов